Часы для изучения временныўх закономерностей развития животных
Изучение времени как естественнонаучной и философской категории имеет долгую, более 25 столетий, историю (Подольный, 1988). Тем не менее задача определения времени в единицах, доступных количественному учету и отражающих качественную специфику изучаемого явления, а также позволяющих выяснить с их помощью временныўе закономерности, остается актуальной и трудно решаемой для многих наук и сегодня. Среди многочисленных определений времени имеется определение, принятое В.И.Вернадским, которое гласит, что время — это дление (Вернадский, 1988). Это определение, как мне представляется, вполне достаточно для наук, изучающих развитие животных. Оно передает основные свойства времени — его непрерывность, необратимость, длительность.
Гораздо труднее найти ответ на вопрос о способах измерения времени или о мере времени. Широко используемые с этой целью единицы астрономического времени (сутки, часы, минуты, секунды) дают очень ограниченную информацию, справедливую в каждом случае только для данного вида организмов и данных конкретных условий (Детлаф, 1986).
Поискам меры времени, которая позволила бы прогнозировать время развития и получить характеристики времени развития, сопоставимые в разных условиях и у разных видов животных, а также выяснить временныўе закономерности развития животных, был посвящен большой ряд исследований, выполненных, главным образом, сотрудниками лаборатории экспериментальной эмбриологии им. Д.П.Филатова в Институте биологии развития им. Н.К.Кольцова РАН (Детлаф Т.А., Детлаф А.А., 1960; 1982; Dettlaff et al., 1961; Детлаф, 1977; Игнатьева, 1979; Dettlaff et al., 1987). В настоящей статье будут кратко изложены основные результаты этих исследований.
§1. Биологическая мера времени
Для решения поставленных задач необходимо было преодолеть большие трудности, состоящие в том, что время развития животных зависит от температуры и у каждого вида имеется своя наследственно обусловленная зависимость скорости развития от температуры. Дополнительную трудность создавало широко распространенное представление (![]() lehrбdek, 1935; Светлов, 1978), согласно которому скорость развития как зачатков разных органов, так и целых зародышей на разных стадиях по-разному зависит от температуры. Продвинуться в решении этих задач нам удалось только после того, как была установлена (Детлаф, 1953; Детлаф Т.А., Детлаф А.А., 1960; Dettlaff Т.А., Dettlaff А.А., 1961) и многократно подтверждена (Детлаф, 1977; Игнатьева, 1979; Dettlaff et al., 1987) ошибочность этого представления и показано, что в зоне оптимальных температур продолжительность разных органогенезов и разных периодов развития зародышей пойкилотермных животных с изменением температуры изменяется пропорционально.
lehrбdek, 1935; Светлов, 1978), согласно которому скорость развития как зачатков разных органов, так и целых зародышей на разных стадиях по-разному зависит от температуры. Продвинуться в решении этих задач нам удалось только после того, как была установлена (Детлаф, 1953; Детлаф Т.А., Детлаф А.А., 1960; Dettlaff Т.А., Dettlaff А.А., 1961) и многократно подтверждена (Детлаф, 1977; Игнатьева, 1979; Dettlaff et al., 1987) ошибочность этого представления и показано, что в зоне оптимальных температур продолжительность разных органогенезов и разных периодов развития зародышей пойкилотермных животных с изменением температуры изменяется пропорционально.
Приведем некоторые из исходных данных, на которых было основано это заключение. На рис.1 представлены графики, выражающие зависимость времени наступления семи последовательных стадий развития зародышей осетра от температуры в часах от осеменения. На рис.2 представлены те же данные, но пересчитанные в числе разных периодов развития: в числе периодов времени между появлением на поверхности яйца борозд первого и второго делений дробления (tn/t0), в числе периодов от осеменения до начала гаструляции (tn/tI) и в долях всего периода зародышевого развития — от осеменения до вылупления зародышей из оболочек (tn/tVII•100%). На рис.3 те же эмпирические данные, определенные в часах, построены в логарифмических координатах. При сравнении рис.1 и 2 легко видеть, что при переходе от характеристики времени в часах к относительной характеристике времени развития числом принятых за единицу времени периодов развития, в зоне оптимальных температур кривые спрямляются и проходят более или менее параллельно оси абсцисс, т.е. продолжительность разных периодов развития зародышей при изменении температуры изменяется пропорционально. Об этом же свидетельствует параллельное и эквидистантное положение кривых на рис.3.
На основании этих исходных данных нами (Детлаф Т.А., Детлаф А.А., 1960; Dettlaff Т.А., Dettlaff А.А., 1961; Детлаф, 1977) был сделан вывод о том, что для измерения продолжительности любого периода зародышевого развития, обозначаемого tn, можно использовать в качестве меры времени продолжительность при той же температуре некоторого принятого за единицу времени периода tx, т.е. можно использовать относительную или безразмерную характеристику продолжительности развития tn/tx. Частное от деления tn на tx показывает, во сколько раз tn больше (или меньше) tx при любой оптимальной температуре. Такой способ определения времени развития позволил нам освободить время от его зависимости от температуры, поскольку величина tn/tx в зоне оптимальных температур при разных температурах одинакова. Он освободил также время от видовой зависимости скорости развития от температуры, т.е. для tn и tx она одинакова.
Наконец, он освободил нас от астрономических единиц времени, которые сокращаются.

Рис.1. Продолжительность развития зародыша осетра от осеменения до начала гаструляции (tI), до конца гаструляции (tII), до смыкания нервных валиков (tIII), до смыкания боковых пластинок в области зачатка сердца (tIV), до начала пульсации сердца (tV), до стадии, когда конец хвоста касается головы (tVI), до вылупления (tVII)
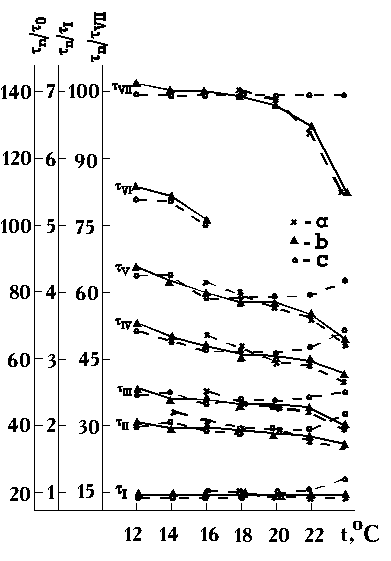
Рис.2. Продолжительность разных периодов развития (tI - tVII) осетра, выраженная в безразмерных единицах: a) в числе периодов одного митотического цикла первых делений дробления (tn/t0), б) — в числе периодов дробления (tn/tI), в) — в процентах от общей продолжительности развития от осеменения до начала вылупления (tn/tVII•100%). По оси ординат — продолжительность периодов развития, выраженная в относительных единицах

Рис.3. Зависимость логарифма продолжительности периодов развития (tI - tVII) от логарифма температуры

Рис.4. Зависимость продолжительности одного митотического цикла в период синхронных делений дробления (t0) у осетра
Для каждого вида животных мерой времени могут служить разные четко засекаемые периоды развития. Однако для того чтобы сравнивать время развития разных животных и выяснять общие для них временныў е закономерности, важно, чтобы принятый за меру времени период развития был равноценен у разных животных. Между тем оказалось, что таких равноценных периодов развития почти не существует вследствие широкого распространения явления гетерохроний: у разных видов животных оплодотворение яйца происходит на разных стадиях мейоза, гаструляция — на разных стадиях дробления, вылупление зародышей из оболочек — на разных стадиях развития и т.п. Исключение представляет, как выяснилось, митотический цикл в период синхронных делений дробления, который проходят в своем развитии большинство пойкилотермных животных — черви, моллюски, членистоногие, иглокожие, рыбы, амфибии, а также птицы (Детлаф, 1977; Ротт, 1980; 1987)1.
Митотический цикл в период синхронных делений дробления характеризуется рядом общих признаков: минимальной продолжительностью, практически одинаковой относительной продолжительностью одноизменных фаз митоза, отсутствием фаз g1 и g2, характерных для полного цикла. В ядрах отсутствуют гранулярные ядрышки и соответственно не происходит синтеза РНК. Цитоплазма бластомеров содержит высокий пул предшественников макромолекулярных синтезов. Наличие всех этих признаков у разных видов животных позволило выбрать в качестве меры времени, сопоставимой у большинства из них, продолжительность одного митотического цикла в период синхронных делений дробления, обозначенную нами символом t0. Ряд исследователей, по предложению Нейфаха, называют ее детлафом (Нейфах, 1961). Продолжительность t0 равна интервалу между появлением одноименных фаз митоза двух последовательных делений дробления.
Для того чтобы использовать t0 в качестве меры времени, надо было для каждого вида животных определить его величину в минутах во всей зоне нерестовых температур и построить графики зависимости t0 от температуры, подобные представленному (рис.4). Продолжительность митотического цикла при разной температуре определяли по продолжительности цикла цитотомии при той же температуре. Предварительно при помощи этнологического изучения было установлено, что митотический цикл у осетровых рыб (Гинзбург, 1959) и у амфибий (Руднева, 1972; Ротт, 1973) равен интервалу между появлением на поверхности яйца борозд I и II делений дробления, а у костистых рыб (Игнатьева, 1979) — половине интервала между появлением борозд II и IV делений дробления (t01) или одной пятой интервала от осеменения до появления борозд IV деления дробления (t02). Графики зависимости t0 от температуры построены для четырех видов осетровых рыб (Детлаф и др., 1993), восьми видов костистых рыб (Игнатьева, Костомарова, 1966; Костомарова, Игнатьева, 1968; Игнатьева, 1979), десяти видов амфибий (Детлаф, 1986; Мазин, Детлаф, 1985) и одного вида морской звезды (Давыдов и др., 1989). Помимо этого величина цикла цитотомии первых делений дробления (предположительно равная митотическому циклу) определена при нескольких температурах у двух видов морских ежей (Бузников, Подмарев, 1975) и у одного вида брюхоногих моллюсков (Мещеряков, 1975).
С помощью этих данных для всех изученных видов животных основное положение о том, что в зоне оптимальных для каждого вида температур продолжительность разных периодов развития с изменением температуры изменяется пропорционально, было подтверждено.
Далее, было показано, что пропорциональность в изменении продолжительности разных периодов развития сохраняется при развитии не только зародышей, но и предличинок, а также и в период созревания ооцитов в яичнике и вне организмов в физиологическом растворе (Детлаф и др., 1993).
Это означает, что с изменением температуры пропорционально изменяется длительность процессов, имеющих самую разную природу и осуществляющихся на разных уровнях организации организма: внутриклеточном (молекулярном и ультраструктурном), клеточном (при делении клеток и их дифференцировке), на уровне морфогенетических движений, процессов индукции и органогенеза. В связи с этим представляется весьма актуальным вопрос о том, каким образом при изменении температуры обеспечивается строгое соответствие изменений временныў х параметров столь различных процессов. Следует предполагать, что оно осуществляется на каких-то глубоких уровнях регуляции, общих для самых разных проявлений жизни и, может быть, входит в функции хаускипинг генов. Мы говорим об этом для того, чтобы привлечь внимание исследователей к изучению этой недавно возникшей проблемы. По мнению Л.Н.Любинской (Детлаф, Любинская, 1987), пропорциональность изменения продолжительности столь разных процессов при изменении температуры может быть описана с помощью методов аффинной геометрии.
Пропорциональность изменений продолжительности разных процессов и разных периодов развития при изменениях температуры в пределах оптимальных для каждого вида пойкилотермных организмов имеет огромное значение. Не будет преувеличением, если мы скажем, что без этой способности пойкилотермные организмы вообще не могли бы существовать в меняющихся условиях внешней среды: если бы разные компоненты комплекса процессов, из которых складывается любой этап развития, изменялись асинхронно, то это приводило бы к возникновению нарушений нормального развития, а на более поздних стадиях — к нарушению нормального функционирования организма. Не случайно, что одной из первых реакций зародышей на приближение к границам оптимальных температур является десинхронизация отдельных процессов развития. В начале она обычно невелика и компенсируется за счет запаса регуляционных возможностей организма (Шмальгаузен, 1938; 1964), но при больших отклонениях температуры от оптимальной выходит за границы регулируемых изменений, и это приводит к развитию уродов и к гибели организма.
Таким образом, пропорциональность изменения продолжительности разных периодов и процессов развития при изменении температуры в пределах биологического оптимума является необходимым условием как целостности развивающегося организма, так и его адаптации к меняющимся условиям среды (Детлаф, 1981; 1985). Она же является надежным основанием для широкого использования метода относительной, безразмерной характеристики продолжительности развития и введения в исследования по биологии развития пойкилотермных животных критерия биологического времени, сопоставимого у разных животных и в разных, но оптимальных для каждого вида температурных условиях.
К сказанному надо прибавить одно важное обстоятельство, которое нельзя забывать, а именно, что относительной мерой времени можно пользоваться только применительно к тем условиям, при которых на изучаемых стадиях развития сохраняется пропорциональность изменения величин tn и t0. Между тем границы оптимальных температур по мере развития могут смещаться (Зотин и др., 1989). Для более поздних стадий они смещаются обычно в сторону несколько более высоких температур, крайние из которых могут уже повреждать зародыш на ранних стадиях. Вследствие этого амплитуда температур, при которых сохраняется пропорциональность изменений tn и t0, в процессе развития может сужаться.
Это в первую очередь касается медленно развивающихся видов, размножающихся осенью, и предличинок весенне-нерестующих рыб.
§2. Использование метода относительной характеристики продолжительности развития для прогнозирования величин tn при разной температуре
Начнем с прогнозирования времени наступления разных стадий и длительности разных периодов развития. Напомним, что поскольку величины tn и t0 в зоне оптимальных для развития температур изменяются пропорционально, то частное от деления tn на t0, определенных в минутах при одной и той же температуре, является величиной постоянной для всей зоны оптимальных температур. Поэтому, определив частное от деления tn на t0 при одной температуре и построив график зависимости t0 от температуры, можно расчетным путем определять величину tn в минутах для любой оптимальной температуры, интересующей исследователя. С этой целью нужно частное от деления tn на t0 умножить на величину t0, снятую с кривой. Такой способ определения tn во много раз сокращает труд исследователя, освобождая его от необходимости получать для каждой температуры новые эмпирические данные. Он позволяет рассчитывать время процессов, трудно доступных для прямого наблюдения, например, происходящих в овариальных ооцитах в полости тела самки или в ядрах делящихся клеток. Кроме того, результаты, полученные методом относительной характеристики продолжительности развития, нередко позволяют уловить ошибки эмпирических наблюдений.
Чтобы пропагандировать предложенный метод относительной характеристики времени развития и облегчить его внедрение в практику научных исследований, в книгах “Объекты биологии развития” (1975) и “Experimental species in developmental biology” (1989) опубликованы данные, полученные для ряда объектов, широко используемых в научных работах и в рыбоводстве. Они содержат таблицы нормального развития (как имевшиеся в литературе, так и специально разработанные для этих изданий), в них рядом с графами “Номер стадии” и “Время наступления в минутах и часах” имеется графа “Время в числе t0”. Для всех этих объектов приведены графики зависимости t0 от температуры.
Кроме того, с помощью метода относительной характеристики продолжительности развития для четырех видов осетровых рыб построены графики, позволяющие рассчитать при разной температуре время инъекции производителям суспензии гипофизов, стимулирующей их созревание, таким образом, чтобы самки созрели в удобное для рыбоводов время. Построены также графики, позволяющие прогнозировать интервал времени, в течение которого следует при разной температуре просматривать инъецированных самок, чтобы получать от них хорошую в рыбоводном отношении икру. Кроме этого, построены графики, позволяющие прогнозировать время наступления стадий, на которых рекомендуется оценивать качество осеменения и типичность развития зародышей. Существенное замедление созревания самок, развития зародышей и предличинок по сравнению с прогнозируемым свидетельствует о недостаточно благоприятных для них условиях среды. Прогнозирование времени созревания самок и развития икры внедрено в практику осетроводства и позволило не только повысить его эффективность, но и облегчило труд рыбоводов, вводя его в оптимальный для них график (Детлаф и др., 1993).
Не меньшее значение имеет возможность прогнозирования времени развития объектов исследования в практике научной работы. В частности, возможно проводить опыты с различного рода воздействиями на разных животных и при разных температурах в течение биологически равнозначного для них времени, измеряемого равным числом или равными долями t0. К сожалению, эта возможность еще не оценена большинством экспериментаторов и мало используется в практике лабораторных исследований.
§3. Временныўе закономерности развития пойкилотермных животных
Остановимся теперь кратко на вопросе о том, какие временныў е закономерности развития удалось выявить с помощью метода относительной безразмерной характеристики продолжительности развития. При этом речь будет идти о времени, заложенном в генетической программе развития каждого данного вида животных, освобожденном от его зависимости от температуры и от наследственно обусловленной зависимости скорости развития от температуры. Эта программа включает определение времени перехода зародышей от одного периода развития к другому и скорости прохождения зародышем последовательных периодов и отдельных морфогенетических процессов. Использование относительной характеристики продолжительности развития позволяет сравнивать их у различных животных. Для сравнения скорости развития у разных животных был введен критерий относительной скорости развития (КОС), равный частному от деления (tn/t0)1 одного вида на (tn/t0)2 другого вида (Детлаф, 1986):
![]() .
.
В тех случаях, когда у сравниваемых видов животных переход от одного периода развития к другому происходит на одной и той же стадии развития, но за меньшее или большее число t0 и КОС у них больше или меньше единицы, мы будем говорить о разной скорости их развития. В тех же случаях, когда у сравниваемых видов переход от одного периода развития к другому запрограммирован на разных стадиях развития, мы имеем дело с гетерохрониями, которые следует отличать от изменения скорости развития. Для выявления гетерохроний достаточно провести тщательные сравнительно-морфологические исследования, а использование относительной характеристики продолжительности развития позволяет датировать их в числе t0, сопоставимом у разных животных и в разных условиях.
Наиболее общей из установленных закономерностей является консерватизм временноўй организации ранних процессов развития у разных животных. Так, продолжительность одноименных фаз митотического цикла в период синхронных делений дроблений у таких далеких объектов, как морской еж, разные виды осетровых и костистых рыб и амфибий, измеряются одинаковыми (или почти одинаковыми) долями t0, т.е. у разных животных занимают одинаковое биологическое время (и это при том, что измеренная минутами длительность их может различаться в десятки раз) (Детлаф, 1962; 1977; Скоблина, 1965; Руднева, 1972; Игнатьева, 1973; 1979). Одинаковым числом t0 измеряется также продолжительность периодов оплодотворения и синхронных делений дробления у зародышей костистых рыб, относящихся к разным семействам (Игнатьева, 1973; Ротт, 1987).
В отличие от самых ранних стадий развития в дальнейшем продолжительность одноименных периодов развития измеряется одинаковым числом t0 обычно только у близкородственных видов, но иногда также у животных, относящихся к разным родам и даже семействам, географически очень удаленным друг от друга. Это установлено на четырех представителях осетровых рыб, относящихся к двум разным родам, двух видах лососевых, двух видах сиговых, разных видах лягушек, разных представителях хвостатых амфибий (Dettlaff et al., 1987).
Установлено, что постоянство относительной скорости развития (КОС =1) сохраняется у родственных видов животных в течение разного времени. Так, у зародышей севрюги, осетра, белуги и стерляди КОС равен единице до середины периода зародышевого развития, а следующие периоды развития зародыши белуги проходят за вcе меньшее число t0, т.е. идут с ускорением (Игумнова, 1975; 1985). Зародыши и предличинки осетра и севрюги развиваются с одинаковой относительной скоростью (КОС =1) по стадии начала жаберного дыхания, но следующий период — от начала жаберного дыхания до начала активного захвата пищи — предличинки осетра проходят за меньшее число t0, чем предличинки севрюги (по данным О.И.Шмальгаузен (Детлаф, 1986)). Когда КОС начинает отклоняться от 1 у разных видов лягушек и у разных представителей хвостатых амфибий — не установлено. Насколько можно судить по таблицам нормального развития, зародыши аксолотля и испанского тритона, относящиеся к разным семействам хвостатых амфибий, одноименные периоды развития проходят за одинаковое число периодов t0 (КОС =1) вплоть до стадии освобождения их из яйцевых оболочек. Для костистых рыб показано, что зародыши двух видов форели развиваются с одинаковой относительной скоростью до стадии 10 пар сомитов, то же относится к двум видам сиговых рыб (пелядь и чир), дальше их развитие прослежено не было (Игнатьева, 1979).
§4. Гетерохронии
При сравнении зародышей разных видов лягушек с зародышами тритонов и аксолотля оказалось, что уже на ранних стадиях обнаруживаются различия в генетической программе временныўх параметров их развития, носящие характер гетерохроний: гаструляция у зародышей тритонов и аксолотля начинается на более ранних стадиях дробления, при меньшем числе t0, чем у лягушек.
Такие же временныўе закономерности развития обнаружены и у костистых рыб (Игнатьева, 1979). После периодов оплодотворения и синхронных делений дробления зародыши разных семейств переходят от бластуляции к гаструляции через время, измеряемое разными числами t0. Гетерохронии в раннем развитии зародышей разных семейств костистых рыб возникают на разных стадиях развития, как и у зародышей хвостатых и бесхвостых амфибий, а именно на тех стадиях, когда для перехода к следующему периоду развития необходимо включение активности генов (Нейфах, 1961; Игнатьева, 1979). Надо надеяться, что дальнейшие исследования умножат наши знания по обсуждаемым вопросам.
§5. Возраст зародышей и его значение
для процессов дифференцировки
Констатация гетерохроний на ранних стадиях развития имеет большое значение для правильной оценки возраста зародышей. Дело в том, что возраст зародышей пойкилотермных животных нельзя определять астрономическими единицами времени, поскольку при разных температурах он был бы при одном и том же уровне дифференцировки разным. Поэтому часто, за неимением других критериев, возраст приравнивают к понятию стадии и считают зародышей разных видов на одинаковой стадии развития равновозрастными. Между тем, как мы уже неоднократно писали (Детлаф, 1956; Dettlaff, 1964), это неправильно, поскольку в результате смещения начала гаструляции на более ранние или поздние стадии дробления у представителей разных систематических групп зародыши на сходных стадиях гаструляции и нейруляции состоят из клеток разных клеточных генераций и содержат разное число клеток. единственно правильным является определение возраста зародыша временем от осеменения, выраженным числом t0. Такое определение позволяет сравнивать возраст зародышей разных животных, находящихся на разных стадиях развития. Возможность оценивать и сравнивать возраст зародышей разных животных и точно определять время их развития впервые позволили ввести параметр времени в сравнительные исследования причинных механизмов индивидуального развития, которыми занимается современная биология развития. Так, например, Г.М.Игнатьева при сравнительном изучении динамики морфогенетических движений в процессе гаструляции у зародышей осетра и аксолотля использовала метод относительной характеристики изменений пространственных и временныў х соотношений между изучаемыми презумптивными зачатками в этот период (время выражено в числе t0 и в долях периода гаструляции, размеры — в угловых градусах и радианах) (Игнатьева, 1979). Это позволило выяснить, каким образом при исходных различиях в топографии и морфогенетических свойствах презумптивных зачатков у сравниваемых видов в процессе гаструляции осуществляется гармоническое сочетание частей и возникает сходное строение зародышей на стадии поздней гаструлы.
Использование понятия “возраст зародыша” имеет большое значение в работах, посвященных сравнительному изучению процессов детерминации, т.е. приобретения клетками зародыша новых свойств, обеспечивающих хотя бы часть пути развития их к конечной цели (образование того или иного органа или части его). Изучение этого процесса представляет одну из центральных задач экспериментальной эмбриологии и, шире, биологии развития. В большой серии работ, посвященных этой проблеме, было обнаружено, что детерминация материала одноименных зачатков органов (плакод органов чувств, нервной пластинки и др.) у разных видов бесхвостых и хвостатых амфибий возникает на разных стадиях развития и по-разному выражена на одинаковых стадиях (Филатов, 1943; Гинзбург, 1959). При этом оказалось, что различия эти коррелируют с различиями в возрасте зародышей на одноименных стадиях развития (Гинзбург, 1989; Детлаф, 1956; Dettlaff, 1964; Dettlaff et al, 1987). Корреляция уровня детерминации с возрастом зародышей ставит вопрос о морфогенетической роли фактора времени. Основания для того, чтобы предполагать существование такой его роли, имелись в литературе и раньше; было обнаружено, что при культивировании эктодермы зародыша в солевом растворе постепенно изменяется ее ответ на индукционные воздействия (Holtfreter, 1938). Однако в те годы вопрос этот еще не мог быть поставлен в полном объеме, в частности из-за отсутствия строгих критериев для оценки возраста зародышей, а также из-за отсутствия экспериментальных подходов к его анализу (Гинзбург, 1989; Dettlaff et al, 1987).
Итак, проблема биологического времени для биологии развития очень важна и очень многогранна. Метод относительной, безразмерной характеристики продолжительности развития открывает широкие возможности для ее изучения, и хочется надеяться, что эти возможности будут использованы в полной мере.
Предложенный нами (Детлаф Т.А., Детлаф А.А., 1960, 1982) безразмерный критерий времени развития животных tn/t0 можно считать оригинальной конструкцией. Она применима для характеристики как времени развития животных, так и для определения их возраста. Сопоставимость этого критерия у разных пойкилотермных животных и в разных оптимальных внешних условиях, а также возможность с его помощью прогнозировать время развития и его изменения в разных условиях и у разных видов животных свидетельствуют о том, что введение этой конструкции правомочно. Более того, она впервые позволила ввести параметр биологического времени в сравнительно-эмбриологические исследования, сделать само время развития объектом изучения и показать его морфогенетическую роль.
Совершенно справедливо высказанное во введении положение о том, что “истинный возраст системы может быть измерен не в астрономической шкале, а в шкале собственного времени системы”, и что для этого должна быть сконструирована и обоснована “собственная шкала”. В настоящей главе возраст в “собственной шкале” измеряется частным от деления tn на t0. Приведены результаты изучения влияния внешних условий (главным образом температуры) на продолжительность и скорость развития. Кроме того, с помощью определения относительной скорости развития ![]() изучен более глубокий, но зависящий от внешних условий уровень регуляции временныўх параметров развития, определяемый геномом зародыша. Воздействия на время развития на генном уровне требуют использования методов генетики и генной инженерии.
изучен более глубокий, но зависящий от внешних условий уровень регуляции временныўх параметров развития, определяемый геномом зародыша. Воздействия на время развития на генном уровне требуют использования методов генетики и генной инженерии.
Длительность разных периодов развития животных выражена числом tn/t0, которое различно для разных стадий развития данного вида животного, но для каждой стадии развития практически постоянно при изменении температуры в границах оптимальной. Это дает основания считать весьма перспективным использование в биологии безразмерных критериев времени.
1Предложенная единица не может быть использована для млекопитающих и некоторых животных с мозаичным типом развития, у которых отсутствует период синхронных делений дробления.
Литература
БУЗНИКОВ Г.А., ПОДМАРЕВ В.И. Морские ежи Strongylocentrotus drobachiensis, S.nudus, S.intermedius. Объекты биологии развития. М., 1975. С.182 - 216.
ВЕРНАДСКИЙ В.И. Философские мысли натуралиста. М., 1988.
ГИНЗБУРГ А.С. Оплодотворение у осетровых рыб. 1. Соединение гамет // Цитология. 1959. Т.1, №5. С.510 - 526.
ГИНЗБУРГ А.С. Детерминация производных эктодермы у разных видов амфибий // Онтогенез. 1989. Т.20, №1. С.47 - 57.
ДАВЫДОВ П.В., ШУБРАВЫЙ О.И., ВАСЕЦКИЙ С.Г. Продолжительность митотического цикла в период синхронных делений дробления (t0) у морской звезды Asterina pectinifera (Mьller et Froschel) // Онтогенез. 1989. Т.20, №1. С.58 - 62.
ДЕТЛАФ Т.А. Зависимость темпа дробления яиц осетровых рыб от температуры // Докл. АН СССР. 1953. Т.91, №3. С.693 - 695.
ДЕТЛАФ Т.А. Видовые различия в формообразовательных свойствах зародышевого материала и смещение гаструляции относительно стадий дробления. Значение соотношения стадий развития и клеточных поколений // Докл. АН СССР. 1956. Т.111, №5. С.1149 - 1152.
ДЕТЛАФ Т.А. Динамика митоза первых делений дробления в яйцах осетра и форели // Ж. общ. биол. 1962. Т.23, №6. С.401 - 409.
ДЕТЛАФ Т.А. Некоторые температурно-временныў е закономерности эмбрионального развития пойкилотермных животных // Проблемы экспериментальной эмбриологии. М. 1977. С.269 - 289.
ДЕТЛАФ Т.А. Адаптация пойкилотермных животных к развитию в условиях колеблющихся температур и проблема целостности развивающегося организма // Онтогенез. 1981. Т.12, №2. С.147 - 152.
ДЕТЛАФ Т.А. Об общих принципах целостности организма в процессе индивидуального развития // Ж. общ. биол. 1985. Т.12, №3. С.227 - 242.
ДЕТЛАФ Т.А. Скорость развития пойкилотермных животных // Ж. общ. биол. 1986. Т.47, №2. С.163 - 172.
ДЕТЛАФ Т.А., ГИНЗБУРГ А.С., Шмальгаузен О.И. Развитие осетровых рыб. Созревание яиц, оплодотворение, развитие зародышей и предличинок. М., 1993.
ДЕТЛАФ Т.А., ДЕТЛАФ А.А. О безразмерных характеристиках продолжительности развития в эмбриологии // Докл. АН СССР. 1960. Т.136, №1. С.199 - 202.
ДЕТЛАФ Т.А., ДЕТЛАФ А.А. Безразмерные критерии как метод количественной характеристики развития животных // Математическая биология развития. М., 1982. С.25 - 39.
ДЕТЛАФ Т.А., ЛЮБИНСКАЯ Л.Н. К вопросу о некоторых механизмах самоорганизации // Самодвижение, самоорганизация, самоуправление. Тезисы докладов межвузовской научной конф. Пермь, 1987.
ЗОТИН А.И., АЛЕКСЕЕВА Т.А., ОЗЕРНЮК А.Д. Метод определения оптимальных условий развития по суммарному потреблению кислорода // Онтогенез. 1989. Т.20, №1. С.103 - 107.
ИГНАТЬЕВА Г.М. Относительная продолжительность процессов карио- и цитомалии в период синхронных делений у карпа и щуки при различных температурах // Онтогенез. 1973. Т.3, №1. С.17 - 21.
ИГНАТЬЕВА Г.М. Ранний эмбриогенез рыб и амфибий. (Сравнительный анализ временныўх закономерностей развития.) М., 1979.
ИГНАТЬЕВА Г.М., КОСТОМАРОВА А.А. Продолжительность митотического цикла в период синхронных делений дробления (t0) и ее зависимость от температуры у зародышей вьюна // Докл. АН СССР. 1966. Т.168, №5. С.1221 - 1224.
ИГУМНОВА Л.В. Временныў е закономерности зародышевого развития белуги // Онтогенез. 1975. Т.6, №6. С.47 - 54.
ИГУМНОВА Л.В. Временныў е закономерности зародышевого развития стерляди Acipenser ruthenus // Онтогенез. 1985. Т.16, №1. С.47 - 54.
КОСТОМАРОВА А.А., ИГНАТЬЕВА Г.М. Соотношение процессов карио- и цитомии в период синхронных делений дробления у вьюна Misgurnus fossilis L. // Докл. АН СССР. 1968. Т.183, №2. С.490 - 492.
МАЗИН А.Л., ДЕТЛАФ Т.А. Зависимость продолжительности одного митотического цикла в период синхронных делений дробления (t0) от температуры у четырех видов Rana и границы оптимальных температур для их размножения и раннего развития // Онтогенез. 1985. Т.16, №4. С.382 - 388.
МЕЩЕРЯКОВ В.Н. Прудовик (Lymnaea stagnalis) // Объекты биологии развития. М., 1975. С.53 - 94.
НЕЙФАХ А.А. Сравнительное радиационное исследование морфогенетической функции ядер в развитии животных // Ж. общ. биол. 1961. Т.22. С.42 - 57.
Объекты биологии развитиЯ. Ред. Т.А.Детлаф. М., 1975.
ПОДОЛЬНЫЙ Р. Спор с тральфадорцами // Знание — сила. 1988. №4. С.39 - 43.
РОТТ Н.Н. Соотношение процессов карио- и цитокинеза при первых делениях дробления у аксолотля (Ambistoma mexicanum Cope) // Онтогенез. 1973. Т.4, №2. С.190 - 196.
РОТТ Н.Н. Клеточные деления в предгаструляционный период развития // Онтогенез. 1980. Т.11, №1. С.3 - 23.
РОТТ Н.Н. Клеточные циклы в раннем эмбриогенезе животных. М., 1987.
РУДНЕВА Т.Б. Продолжительность карио- и цитомии в период II — IV делений дробления у шпорцевой лягушки Xenopus laevis // Онтогенез. 1972. Т.3, №6. С.622- 626.
СВЕТЛОВ П.Г. Физиология (механика) развития. Т.2. Л., 1978. С.188 - 196.
СКОБЛИНА М.Н. Безразмерная характеристика продолжительности фаз митоза первых делений дробления у аксолотля // Докл. АН СССР. 1965. т.160, №3. С.700 - 703.
ФИЛАТОВ Д.П. Механика развития как метод изучения некоторых вопросов эволюции // Ж. общ. биол. 1943. Т.4. С.28 - 64.
ШМАЛЬГАУЗЕН И.И. Организм как целое в индивидуальном и историческом развитии. М., 1938.
ШМАЛЬГАУЗЕН И.И. Регуляция формообразования в индивидуальном развитии. М., 1964.
![]() HRБDEK J. Temperature and living matter. Protoplasma Monographien Bd. 8. Berlin, 1935.
HRБDEK J. Temperature and living matter. Protoplasma Monographien Bd. 8. Berlin, 1935.
DETТLAFF T.A. Cell divisions, duration of interkinetic states and differentiation in early stages of embrionic development // Adv. Morphogen. 1964. №3. P.323 - 362.
DEТTLAFF T.A., DEТTLAFF A.A. On relative dimensionsless characteristics of the development duration in embriology // Arch. Biol. (Liege). 1961. V.72. P.1 - 16.
DETТLAFF T.A., IGNATIEVA G.M., VASSETZKY S.G. The problem of time in developmental biology: its study by the use of relative characteristics of developmental duration // Sov. Sci. Rev. Ser. F.Physiol. Gener. Biol. 1987. V.1. P.1 - 88.
Experimantal species for developmental biology. Dettlaff T.A., Vassetzry S.G. (Eds.). Plenum Press. 1989.
HOLTFRETER J. Differenzierungspotenzen isolierter teile der urodelengastrula // W. Roux’ Arch. Entwicklungamech. Organismen. в.138. s.522 - 656.