Проблемы окружающей среды (обзорная информация ВИНИТИ), № 9, 2002.
© П.В.Фурсова, А.П.Левич
МАТЕМАТИЧЕСКОЕ МОДЕЛИРОВАНИЕ В ЭКОЛОГИИ СООБЩЕСТВ
Обзор литературы
П.В.Фурсова, А.П.Левич
Аннотация
Обзор литературы посвящен математическому моделированию в экологии сообществ, в частности, моделированию сообществ фитопланктона и бактериальных сообществ.
В первой части содержится информация об общих принципах и задачах моделирования. Кратко представлены классы моделей, основные этапы их построения, перечислены математические инструменты, привлекаемые на разных шагах моделирования.
Во второй части обзора представлены математические методы построения моделей: дифференциальные уравнения, вариацонное исчисление, клеточные автоматы, нейронный сети, "организменные" модели.
Третья часть посвящена подробному изучению моделей фитопланктонных и микробиологических сообществ, построенных с использованием дифференциальных уравнений. Приведены примеры типов уравнений, применяемых в данных моделях. Описана концепция лимитирующих факторов и способы ее математической формализации. Представлено моделирование конкуренции за ресурсы, в частности, моделирование совместного потребления ресурсов питания организмами одного трофического уровня. Как результат конкуренции особо рассматривается перестройка структуры фитопланктонного сообщества, а также вопрос о конкурентном исключении видов. Описана теория межвидовой "борьбы", в которой аналитическое исследование уравнений заменено графическим подходом. Упомянуто моделирование влияния миграции видов на устойчивость сообщества. Представлены модели, допускающие управление ростом и урожаем.
В четвертой части представлен обзор ряда экстремальных принципов, применяемых в биологии, и основанных на этих принципах методов вариационного моделирования. Особое внмание уделено экологическим приложениям к развитию “формализма Джейнса”, состоящего в условной оптимизации энтропийных целевых функций.
Обзор предназначен для специалистов по математическому моделированию, теории систем, для экологов и специалистов по охране природы. Обзор доступен аспирантам и студентам соответствующих специализаций.
Введение
Математическое моделирование в экологии сообществ – достаточно обширная область исследования и по выбору объектов моделирования, и по набору методов, и по спектру решаемых задач. Предлагаемый читателю обзор не претендует на охват всех аспектов моделирования. Внимание авторов обращено на два класса методов: моделирование с помощью дифференциальных уравнений и методы, основывающиеся на экстремальных принципах биологии. Если примеры вариационных моделей относятся к довольно широкому кругу растительных и животных сообществ, то для подходов, основанных на дифференциальных уравнениях, в виду обширности материала внимание сконцентрировано на моделировании сообществ микроорганизмов.
Модели каждого из методов, безусловно, обладают своими достоинствами и недостатками. Так, дифференциальные или разностные уравнения позволяют описывать динамику процессов в режиме реального времени, тогда как вариационные методы, как правило, предсказывают лишь конечное стационарное состояние сообщества. Но на пути имитаций с помощью уравнений возникают трудности как принципиального, так и технического характера. Принципиальная трудность состоит в том, что не существует систематических правил вывода самих уравнений. Процедуры их составления основываются на полуэмпирических закономерностях, правдоподобных рассуждениях, аналогиях и искусстве модельера. Технические трудности связаны с высокой размерностью задач по моделированию сообществ. Для существенно многовидовых сообществ, потребляющих многочисленные ресурсы, требуется подбор сотен коэффициентов и анализ систем из десятков уравнений. (Если изучается сообщество из w групп организмов, потребляющих m ресурсов, то соответствующая система дифференциальных уравнений должна содержать, по крайней мере, w + mw + m уравнений с 2w + 4mw параметрами, требующими идентификации.) Обычные приемы снижения числа переменных – их агрегирование или учет только доминирующих групп организмов – непригодны во многих задачах экологии. С течением времени существенную роль начинают играть редкие и малочисленные виды, которые, тем самым, следует включать в число переменных на начальных этапах моделирования. Агрегация переменных может нивелировать результаты управления функционированием сообществ. При работе с системами из десятков и более дифференциальных уравнений оказывается, что проследить причинные связи (для отладки, исключения ошибок, интерпретаций) в системе уравнений также сложно, как и в реальной экосистеме. В конце концов, оказывается, что мы не можем узнать, чему обязаны полученными результатами: реальному положению вещей, ошибкам в исходных данных, недочетам алгоритма или еще чему-либо. Модели, основанные на экстремальных принципах, как правило, преодолевают "проклятие размерности", но сохраняют произвол в выборе самих исходных принципов.
В обзоре для каждого из подходов приводится исходная формулировка модели и примеры ряда получаемых в модели биологически интерпретируемых результатов.
Авторы благодарны В.Л.Алексееву за ряд предложенных им материалов, включенных в обзор.
1. Общие принципы и задачи моделирования
В зависимости от цели моделирования, можно выделить два типа моделей: дескриптивные модели и модели поведения (Страшкраба, Гнаук, 1989).
Дескриптивная модель позволяет получить информацию о взаимосвязях между наиболее важными переменными экосистемы. Реализуется такой тип модели методами стохастического моделирования, основанного на инструментах теории вероятностей и математической статистики. Разделяют статические методы, не учитывающие время в качестве переменной (простая и множественная линейная и нелинейная корреляция и регрессия; дисперсионный, дискриминантный и факторный виды анализа, методы оценки параметров), и динамические методы, которые учитывают временную переменную (анализ Фурье, корреляционный и спектральный анализ, весовые и передаточные функции). В отечественной литературе подобные модели, представляющие собой регрессионные и другие эмпирически установленные количественные зависимости, не претендующие на раскрытие механизма описываемого процесса, получили название описательных (Ризниченко, Рубин, 1993).
Модели поведения описывают системы во время переходного периода от одного состояния к другому (Страшкраба, Гнаук, 1989). Для осуществления этой категории моделей изучают: 1) структуру сигналов на входе и выходе системы; 2) реакцию системы на особые проверочные сигналы; 3) внутреннюю структуру системы. Последний пункт реализуется аналитическим моделированием, в основе которого лежат дифференциальные уравнения, описывающие причинно-следственные связи в экосистеме. Первым этапом аналитического моделирования является формирование концепции модели и составление уравнений, описывающих поведение системы, при этом происходит упрощение реальности, которое, однако, не влияет на наиболее существенные свойства реальной системы. Затем идет параметризация, т.е. определение количественных значений параметров. Осуществление этой задачи возможно тремя способами: 1) получением предварительных оценок значений параметров на основе наблюдений; 2) нахождением комбинаций параметров, отвечающих моделируемой ситуации, базирующимся на методах оптимизации параметров; 3) оценкой роли параметров модели с помощью анализа чувствительности, целью которого является определение того, как модель реагирует на изменение значений параметров и, как следствие, того, насколько правильно оценены параметры. Следующий шаг аналитического моделирования – имитация, т.е. получение с помощью ЭВМ решения модельных уравнений при фиксированных значениях параметров и начальных условиях. И, наконец, испытание модели или, другими словами, сравнение ее выходных параметров с выходными данными системы. Различают два способа испытания: 1) проверка (качественное или количественное сравнение данных, полученных в результате моделирования, с действительными значениями); 2) проверка значимости модели (проведение экспериментов для изучения поведения модели и системы с целью обнаружения их сходства, а также для сравнения тенденций поведения модели и системы). Выделяется также адаптивное моделирование, при котором происходит автоматическая адаптация модели к системе с помощью ЭВМ.
Классификация математических моделей биологических продукционных процессов была предложена в книге Г.Ю.Ризниченко и А.Б.Рубина (1993). Различают три класса: 1) описательные модели; 2) качественные модели (выясняющие динамический механизм изучаемого процесса, способные воспроизвести наблюдаемые динамические эффекты в поведении системы); 3) имитационные модели конкретных сложных систем, учитывающие всю имеющуюся информацию об объекте (и позволяющие прогнозировать поведение систем или решать оптимизационные задачи их эксплуатации). Особое значение придается именно последнему классу моделей, поскольку он оказывается полезным для практических целей. Кратко можно выделить следующие основные этапы построения имитационной модели (Ризниченко, Рубин, 1993):
Однако практика показала, что попытки детального описания многокомпонентных систем приводит к проблеме “проклятия размерности”, когда практически невозможно корректное построение и идентификация математической модели из-за использования чрезмерно большого количества неточно определенных параметров по сравнению с имеющейся экспериментальной информацией (Алексеев и др., 1992). В такой ситуации необходимо упрощение модели, например, за счет отбрасывания блоков или функциональных связей с второстепенным значением, выделения наиболее важных составляющих, определения быстрых и медленных переменных и замены части из них постоянными величинами или параметрическими зависимостями.
2. Краткое представление методов, применяемых для построения моделей
2.1. Дифференциальные уравнения. Для описания экологических сообществ привлекают методы из самых разных областей математического знания. Самое широкое распространение получил подход, основывающийся на аппарате дифференциального исчисления. Дифференциальные уравнения позволяют описывать динамику численности (биомассы) каждой популяции, входящей в изучаемую систему. В общем виде можно записать зависимость
![]() (2.1)
(2.1)
где w – число видов в сообществе, xi – численности i-го вида, t – время. Если сделать предположения о стационарности среды и изолированности сообщества (т.е. исчезнувший вид не может возникнуть вновь, а это значит, что ![]() ), то уравнения (2.1) можно записать в виде (Абросов, Ковров, 1977)
), то уравнения (2.1) можно записать в виде (Абросов, Ковров, 1977)
![]()
Дальнейшее теоретическое исследование в большинстве случаев проводится для конкретного вида функций ![]() .
.
2.2. Вариационное исчисление. Принципиально иным является метод моделирования, основанный на применении экстремальных принципов. Согласно им в реальности осуществляются лишь некоторые состояния системы, а именно, состояния с экстремальным значением числовой функции или функционала, называемых “целевой функцией”, которая определяет развитие природной системы. Широкое применение экстремальные принципы получили в физике, механике, термодинамике, экономике, теории управления. В биологии вопрос о “целевой функции” стал более популярен с распространением эволюционного мышления в противовес статическому видению мира (Wilhelm, Brüggemann, 2000). По Дарвину “выживает самый приспособленный”, но ответ на вопрос “Что является самым приспособленным?” требует адекватной “функции приспособленности”. Можно перечислить следующие предлагавшиеся целевые функции: максимальный поток энергии через систему (Lotka, 1922), максимум "силы" (Odum, Pinkerton, 1955), максимальное рассеяние энергии (Ulanowicz, Hannon, 1987; Schnieder, Kay, 1994; Mauersberger, 1996), запасание "максимальной эксергии" (Jørgensen, 1997), асценденция (Ulanowicz, 1986), индекс зрелости (Pérez-España, Arreguin-Sánchez, 1999), максимизация косвенных эффектов (Patten, 1986; 1995), максимизация биомассы (Margalef, 1968), специальная иерархическая организация (O’Neill et al., 1986), максимум устойчивости органического вещества (Whittaker, Woodwell, 1971), максимизация обобщенной энтропии сообщества или его разнообразия (Левич, 1980), минимизация диссипации энергии и "наискорейший спуск" (А.И.Зотин, А.А.Зотин, 1999). Более подробное описание некоторых целевых функций, экстремальных принципов и их реализаций в математических моделях будет приведено в разделе 4.
2.3. Клеточные автоматы. В моделировании динамики растительного покрова успешно применяют теорию клеточных автоматов. Клеточные автоматы – динамические модели с дискретным временем, пространством и состояниями. Простой клеточный автомат определяется решеткой L, пространством состояний Q, шаблоном соседей δ и функцией местных переходов f. Каждая клетка решетки L может находиться в состоянии из пространства Q. Клетки могут соединяться различными способами, в простейшем случае они образуют квадратную или шестиугольную решетку. Клетки могут менять свое состояние в шаги дискретного времени, обычно они это делают синхронно. Судьба клетки зависит от окружающих ее соседей (соседи первого рода – центральная клетка и четыре примыкающих, соседи второго рода – центральная клетка и 8 примыкающих) и соответствующей функции перехода. Правила перехода определяются формулой
![]()
где ![]() – состояние клетки s в момент времени t; τ – πанг соседних клеток; f – функция местного перехода. Набор величин
– состояние клетки s в момент времени t; τ – πанг соседних клеток; f – функция местного перехода. Набор величин ![]() называется конфигурацией клеточного автомата в момент времени t (I – множество индексов клеток) (Balzter et al, 1998).
называется конфигурацией клеточного автомата в момент времени t (I – множество индексов клеток) (Balzter et al, 1998).
Клеточный автомат с соседями второго рода можно описывать с помощью марковских цепей (Logofet, Lesnaya, 2000). Пусть ![]() – дискретные случайные величины в определенные равноудаленные моменты времени t
– дискретные случайные величины в определенные равноудаленные моменты времени t![]() T, где
T, где ![]() – пространство параметров марковской цепи. Переходные вероятности от одного состояния к другому записываются как
– пространство параметров марковской цепи. Переходные вероятности от одного состояния к другому записываются как ![]() (вероятность того, что переменная
(вероятность того, что переменная ![]() будет в состоянии in+1 в момент времени tn+1 при условии, что переменная
будет в состоянии in+1 в момент времени tn+1 при условии, что переменная ![]() была в состоянии in на предыдущем шаге). Для марковских цепей определяющим является свойство
была в состоянии in на предыдущем шаге). Для марковских цепей определяющим является свойство
![]()
Набор индексов марковской цепи можно рассматривать как пространственный или обозначающий специальные стадии. Распространяя индекс t на временное и пространственное измерение, можно описать клеточный автомат с соседями второго рода. В этом случае пространственная зависимость выражается в том, что будущее клетки s зависит от окружения δs, описываемого простой марковской цепью. По времени будущее клетки зависит только от последнего состояния.
Применение теории клеточных автоматов для моделирования динамики трех видов растительного покрова можно найти в работе Х.Балцтера с соавторами (Balzter et al, 1998), для моделирования сукцессии лесов – в работе Д.О.Логофета и Е.В.Лесной (Logofet, Lesnaya, 2000).
2.4. Нейронные сети. В последнее десятилетие наблюдается всплеск в области исследования и применения искусственных нейронных сетей. Этот метод уже получил распространение в и химических исследованиях, в медицине, молекулярной биологии, экологии (моделирование пространственной динамики рыб, предсказание воспроизводства фитопланктона, разнообразия рыб и т. д.), в исследованиях по распознаванию образов и речи. Обзор основных типов искусственных нейронных сетей представлен в работе С.Лека и Дж.Гуегана (Lek, Guegan, 1999). В зависимости от поставленной задачи (обобщение, оптимизация, управление, предсказание, редукция данных, и т. д.) рассматривают и применяют различные виды нейронных сетей. В настоящее время наиболее активно используются два их типа:
Модель первого типа применяли для моделирования эвтрофикации (Karul et al., 2000). Использовалась сеть, состоящая их трех слоев. Входящий слой состоял из 9-ти нейронов (PO4, NO3, PH, температура воды и другие показатели). Выходящий слой состоял из одного нейрона (хлорофилл А). Для работы с нейронной сетью были использованы данные по озерам Моган и Эймир и водохранилищу Кебан Дам. В работе предъявлено хорошее соответствие данных наблюдений и результатов расчетов с помощью нейронной сети. Однако для сети, в которой в качестве выходящих параметров были использованы три вида фитопланктона, результаты не были столь успешны. Авторы объяснили это количеством и природой доступных им данных по фитопланктону.
2.5. "Организменные" модели. С 1970 года в экологическом моделировании активно используется подход, в рамках которого основным объектом модели является индивид – individual-based models. Индивид рассматривают как уникальную, дискретную единицу, у которой есть по крайней мере еще одна черта в дополнении к возрасту, которая меняется в течение жизненного цикла: вес, ранг в социальной иерархии и т. п. Модели, “основанные на индивиде”, строят снизу вверх, начиная с “частей” (индивидов) системы (популяции). Целью исследования часто становится понимание того, каким образом свойства системы возникают из взаимодействия между частями (Grimm, 1999).
3. Модели, основанные на дифференциальных уравнениях
3.1. Примеры уравнений. В разделе о моделировании с помощью дифференциальных уравнений в первую очередь рассматриваются модели фитопланктонных и микробиологических сообществ.
3.1.1. Моделирование сообществ фитопланктона. Традиционный путь изучения сообществ микроорганизмов заключается в моделировании непрерывных культур. Общее уравнение, описывающее кинетику концентрации клеток в таком процессе, имеет вид
![]()
где x – концентрация клеток в культиваторе, ![]() – функция, описывающая размножение популяции, d – скорость вымывания. Скорость размножения может зависеть от концентрации клеток, концентрации субстрата s, температуры, pH среды и прочих факторов (Ризниченко, Рубин, 1993).
– функция, описывающая размножение популяции, d – скорость вымывания. Скорость размножения может зависеть от концентрации клеток, концентрации субстрата s, температуры, pH среды и прочих факторов (Ризниченко, Рубин, 1993).
В микробиологических системах, как правило, скорость роста лимитируется концентрацией субстрата, что отражается зависимостью, предложенной Ж.Моно (Monod, 1942):
![]()
где ![]() – максимальная скорость роста организмов при данных условиях,
– максимальная скорость роста организмов при данных условиях, ![]() – видоспецифическая константа, численно равная концентрации субстрата, при которой скорость роста культуры равна половине максимальной (константа полунасыщения).
– видоспецифическая константа, численно равная концентрации субстрата, при которой скорость роста культуры равна половине максимальной (константа полунасыщения).
При моделировании динамики фитопланктона важную роль играет учет влияния уровня освещенности на скорость роста. В книге С.Йоргенсена (1985) описаны некоторые виды уравнений, применяемых в моделях. Зависимость между скоростью роста (скоростью первичного продуцирования) и освещенностью может быть описана уравнением Михаэлиса-Ментен
![]()
здесь I – освещенность, KI – константа полунасыщения по освещенности.
При освещенности выше пороговой (IH), начиная с которой происходит угнетение фотосинтеза, можно записать следующую зависимость:
![]()
Значения всех констант (![]() ) зависят от адаптации к освещенности и температуры. Зависимость KI от температуры, как правило, линейна:
) зависят от адаптации к освещенности и температуры. Зависимость KI от температуры, как правило, линейна: ![]() ; am, KI0 – константы, KI0 – константа полунасыщения по освещенности в нулевой точке температурной шкалы. Зависимость IH от температуры так же может быть выражена линейной функцией:
; am, KI0 – константы, KI0 – константа полунасыщения по освещенности в нулевой точке температурной шкалы. Зависимость IH от температуры так же может быть выражена линейной функцией: ![]() ; IH0 – пороговая освещенность в нулевой точке температурной шкалы, aH – константа.
; IH0 – пороговая освещенность в нулевой точке температурной шкалы, aH – константа.
Как уже было сказано, скорость роста зависит от концентрации биогенных веществ. Для фитопланктона элементами, способными лимитировать рост, могут являться, например, азот, фосфор и углерод. Возможные способы отражения этого факта в работе С.Йоргенсена описаны следующим образом
![]() ,
,
где P, N, C – концентрация растворенного фосфора, азота и углерода; KP, KN, KС – соответствующие константы полунасыщения. Возможно также использование средней величины лимитирующих факторов: 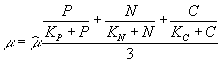 .
.
В работах Н.С.Абросова с коллегами (Абросов, Ковров, 1977; Абросов, Боголюбов, 1988; Абросов, 1999) была предложена следующая модель динамики экосистемы. Рассматривают организмы одного трофического уровня, которые существуют за счет совместного потребления ресурсов питания. Пусть j ij(s1,...,sm) – скорость потребления единицей биомассы (особью) вида i ресурса j; m i = m i(s1,...,sm) – удельная скорость прироста биомассы (плотности) i-го вида; s = (s1,...,sm) – вектор концентрации ресурсов в среде; di – удельная скорость элиминации организмов i-го вида; Аj – скорость поступления j-го субстрата извне на единицу объема экосистемы; Di – удельная скорость удаления ресурса в открытой системе. Тогда динамика системы описывается уравнениями
 (3.1)
(3.1)
Модель, описывающая культивируемое в хемостате сообщество, является частным случаем модели (3.1) при условиях di = Dj = D, ![]() = Aj/D, где D – скорость разведения культуры,
= Aj/D, где D – скорость разведения культуры, ![]() – концентрация j-го субстрата в поступающей в реактор питательной среде, и задается системой уравнений
– концентрация j-го субстрата в поступающей в реактор питательной среде, и задается системой уравнений
![]()
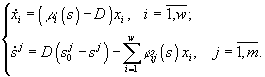
(Хемостат – непрерывный культиватор, в котором за счет саморегулирующх механизмов популяции поддерживается постоянная плотность популяции.)
В случае взаимозаменяемых ресурсов зависимость m i(s) описывается выражением
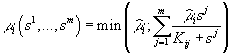
или его линейной аппроксимацией
 ,
,
где b
ij – элементы матрицы приспособленности: 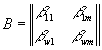 , здесь b
ij = cijYij, где cij – удельная скорость потребления j-го субстрата i-м видом организма, а Yij – экономический коэффициент использования j-го вещества i-м видом (Абросов, Ковров, 1977).
, здесь b
ij = cijYij, где cij – удельная скорость потребления j-го субстрата i-м видом организма, а Yij – экономический коэффициент использования j-го вещества i-м видом (Абросов, Ковров, 1977).
Если ресурсы взаимонезаменяемы, то
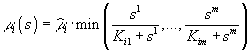
или
![]() .
.
В приведенных выше формулах символ ![]() обозначает максимально возможную удельную скорость роста организмов i-го вида; Kij – константа полунасыщения роста i-го вида на j-м субстрате.
обозначает максимально возможную удельную скорость роста организмов i-го вида; Kij – константа полунасыщения роста i-го вида на j-м субстрате.
В моделях, использующих дифференциальные уравнения, возможен учет лимитирования роста световой энергией. В этом случае предлагается использовать следующую зависимость:
![]()
здесь xi – концентрация клеток i-го вида; μi – удельная скорость роста i-го вида; ![]() – максимально возможная удельная скорость роста организмов i-го вида;
– максимально возможная удельная скорость роста организмов i-го вида; ![]() – процентное содержание хлорофилла в биомассе; li – коэффициент поглощения света i-м видом;
– процентное содержание хлорофилла в биомассе; li – коэффициент поглощения света i-м видом; ![]() – КПД фотосинтеза i-го вида клеток; Yi – величина, обратная калорийности биомассы i-го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора (приведено выражение для удельной скорости μi в случае лимитирования совместного роста двух видов только энергией света) (Абросов, Ковров, 1977).
– КПД фотосинтеза i-го вида клеток; Yi – величина, обратная калорийности биомассы i-го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора (приведено выражение для удельной скорости μi в случае лимитирования совместного роста двух видов только энергией света) (Абросов, Ковров, 1977).
А.Я.Болсуновским (1999) была предложена следующая математическая модель динамики компонентов сообщества из двух видов (спирулина и хлорелла), рост которых лимитируется интенсивностью света всей области ФАР
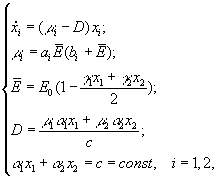
где xi – концентрация клеточного хлорофилла; μi – удельные скорости роста; ai, bi – константы уравнения Михаэлиса-Ментен; ![]() – средняя освещенность для всего сообщества; E0 – поверхностная освещенность; D – скорость протока.
– средняя освещенность для всего сообщества; E0 – поверхностная освещенность; D – скорость протока.
Кроме того, в моделях возможно отразить эффекты метаболического воздействия, как, например, это было сделано Ю.А.Домбровским с соавторами (1990). Скорость роста фитопланктона i -го вида описывалась выражением
![]() ,
,
где x1, x2, s – концентрации двух видов фитопланктона и минерального вещества, выраженные в единицах лимитирующего биогенного вещества; ![]() – коэффициенты метаболизма; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения;
– коэффициенты метаболизма; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения; ![]() – эмпирический коэффициент ингибирования i-го вида j-м.
– эмпирический коэффициент ингибирования i-го вида j-м.
Как уже указывалось выше, при моделировании динамики биомассы фитопланктона, чаще всего используют уравнение Моно (Monod, 1942), описывающее зависимость удельной скорости роста популяции μ от концентрации s лимитирующего рост субстрата
![]() , (3.2)
, (3.2)
где ![]() – максимальная скорость роста; Ks – константа полунасыщения при лимитировании данным субстратом. (В общем случае
– максимальная скорость роста; Ks – константа полунасыщения при лимитировании данным субстратом. (В общем случае ![]() и Ks зависят от вида фитопланктона.) Теоретическое объяснение указанной зависимости обычно основывается на представлениях биохимической кинетики, связанных с концепцией “узкого места” метаболизма (Алексеев и др., 1992). Живая клетка представляет собой строго сбалансированную систему ферментативных реакций, при этом общая скорость прироста биомассы
и Ks зависят от вида фитопланктона.) Теоретическое объяснение указанной зависимости обычно основывается на представлениях биохимической кинетики, связанных с концепцией “узкого места” метаболизма (Алексеев и др., 1992). Живая клетка представляет собой строго сбалансированную систему ферментативных реакций, при этом общая скорость прироста биомассы ![]() предполагается пропорциональной скорости реакции, которая ограничена недостатком субстрата в среде. В простейшем случае зависимость скорости ферментативной реакции от концентрации субстрата описывается формулой Михаэлиса-Ментен. Поскольку в формуле Михаэлиса-Ментен константы
предполагается пропорциональной скорости реакции, которая ограничена недостатком субстрата в среде. В простейшем случае зависимость скорости ферментативной реакции от концентрации субстрата описывается формулой Михаэлиса-Ментен. Поскольку в формуле Михаэлиса-Ментен константы ![]() и Ks есть функции от скоростей прямой и обратной ферментативной реакции, то постоянство
и Ks есть функции от скоростей прямой и обратной ферментативной реакции, то постоянство ![]() и Ks предполагает постоянство этих скоростей. Таким образом, в основе использования зависимости Моно лежит неявное предположение об инвариантности во времени соотношения элементов в клетке (так как только при постоянстве состава клетки могут оставаться постоянными скорости ферментативных реакций). Кроме того, при подходе, основанном на зависимости Моно, процессы поглощения питательных веществ и роста фитопланктона не различаются.
и Ks предполагает постоянство этих скоростей. Таким образом, в основе использования зависимости Моно лежит неявное предположение об инвариантности во времени соотношения элементов в клетке (так как только при постоянстве состава клетки могут оставаться постоянными скорости ферментативных реакций). Кроме того, при подходе, основанном на зависимости Моно, процессы поглощения питательных веществ и роста фитопланктона не различаются.
Существуют, однако, данные, что представление о постоянстве клеточного состава не является верным. Как показали эксперименты, содержание фосфора в одноклеточных водорослях одного вида может в зависимости от внешних условий различаться в 10-30 раз (Финенко, Крупаткина-Акинина, 1974; Крупаткина, 1978), азота – до 5 раз (Sjoberg, 1980), углерода до 12 раз (Dauta, 1982а). Крайне вариабельно также соотношение элементов в клетках. Так С.Йоргенсен (Jørgensen, 1979) для естественных озерных сообществ пресноводных микроводорослей приводит диапазон отклонений N:P от 4,1 до 291. Б.Н.Гутельмахер и Н.А.Петрова (Gutelmacher, Petrova, 1982), непосредственными авторадиографическими измерениями обнаружили в пресноводном озерном фитопланктоне изменчивость отношений запасов углерода к запасам фосфора в клетках от 3,3 до 500.
Поскольку предположения, лежащие в основе зависимости Моно, часто нарушаются, естественно ожидать, что в экспериментах возможны ситуации, которые принципиально не могут быть описаны моделью вида Моно. И такие ситуации действительно наблюдаются. Неоднократно описаны случаи, когда при почти полном отсутствии минеральных веществ в среде (s = 0 в уравнении Моно) фитопланктон, тем не менее, продуцирует новое вещество (m ¹ 0) — явление, как пишет Маккарти, на уровне парадокса – “что-то из ничего” (something from nothing). Другим примером является так называемый феномен избыточного потребления, который известен уже давно (Ketchum, 1939) и красочно описан Э.Кензлером и Б.Кетчумом (Kuenzler, Ketchum, 1962). В их экспериментах с морской диатомеей Phaeodactylum tricornutum в ситуации, когда клетки водоросли были помещены в новую среду, наблюдалось, что сначала практически весь фосфор был потреблен клетками из среды, и только потом клетки начали делиться.
Таким образом, процессы поглощения минеральных веществ из среды клетками фитопланктона и его рост – существенно независимые процессы. И, следовательно, модели, игнорирующие этот факт, не отразят, по крайней мере, три явления, наблюдаемые в природе (Кучай, 1985):
Основными величинами при описании зависимости скорости роста от клеточных запасов биогенных элементов являются непосредственно содержание элемента в клетке q (“клеточная квота”), а также его минимальное содержание в клетке qmin, при котором скорость роста μ = 0.
Развитие концепции клеточной квоты в исследованиях различных авторов описано в работе Л.А.Кучай (1985). Концепция клеточной квоты берет начало с работ М.Друпа, который предложил зависимость удельной скорости роста от содержания вещества в клетке в виде (Droop, 1973)
![]() . (3.3)
. (3.3)
Формула (3.3) упрощается в связи с предположением, подтвержденным экспериментально Г.Ри (Rhee, 1978), что константа полунасыщения и минимальное клеточное содержание соответствующего биогена равны, и принимает вид
![]() .
.
Дж.Фухсом (Fuhs, 1969) в эксперименте на двух видах диатомовых продемонстрирована связь скорости роста водорослей с клеточной концентрацией фосфора в виде ![]() (Обозначения те же, что и в формулах (3.2), (3.3).)
(Обозначения те же, что и в формулах (3.2), (3.3).)
В.Бьерман (Bierman, 1976) в модели внутригодовой сукцессии фитопланктона (четыре группы водорослей, три биогенных элемента – азот, фосфор, кремний) выражал зависимость роста от клеточного фосфора формулой  , от клеточного азота – формулой Друпа (3.3), а от клеточного кремния – формулой Моно (3.2).
, от клеточного азота – формулой Друпа (3.3), а от клеточного кремния – формулой Моно (3.2).
Кроме минимального содержания элемента qmin в клетке, важной характеристикой вида является величина qmax – максимальное значение элемента питания, которое может накопиться в клетке. Можно привести несколько примеров использования величины qmax в формулах для скорости роста клеток (Кучай, 1985). C.Сьеберг (Sjöberg, 1980) в модели весеннего цветения диатомовых на Балтике применял зависимость
![]()
А.Дота (Dauta, 1982б) в модели поликультуры пресноводных водорослей использовал выражение
![]()
На непостоянство стехиометрических соотношений углерода, азота и фосфора в составе фитопланктона указывал С.Йоргенсен (1985). Он отмечал, что, как показали лабораторные опыты, развитие фитопланктона происходит в два этапа: на первом идет поглощение биогенных веществ, а на втором – клеточное деление, регулируемое внутриклеточной концентрацией ресурсов. С.Йоргенсен (Jørgensen, 1976) использует следующую систему уравнений
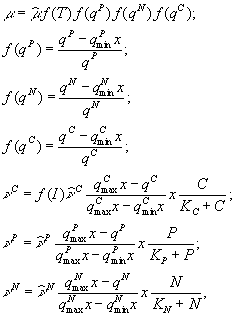
где ![]() – внутриклеточные концентрации биогенных элементов (соответственно, фосфора, азота и углерода), выраженные в мг на 1 л воды;
– внутриклеточные концентрации биогенных элементов (соответственно, фосфора, азота и углерода), выраженные в мг на 1 л воды; ![]() – минимальные и максимальные значения содержания трех биогенных веществ в составе фитопланктона; x – концентрация фитопланктона;
– минимальные и максимальные значения содержания трех биогенных веществ в составе фитопланктона; x – концентрация фитопланктона; ![]() – скорости поглощения углерода, фосфора, азота;
– скорости поглощения углерода, фосфора, азота; ![]() – соответствующие максимальные скорости поглощения; C, P, N – концентрации биогенных элементов в окружающей среде;
– соответствующие максимальные скорости поглощения; C, P, N – концентрации биогенных элементов в окружающей среде; ![]() – функции, характеризующие зависимости интенсивности фотосинтеза от освещенности и температуры.
– функции, характеризующие зависимости интенсивности фотосинтеза от освещенности и температуры.
Существуют некоторые данные, которые позволяют сравнить результаты моделирования с использованием концепции клеточной квоты и без ее использования. Л.А.Кучай (Кучай, 1985) специально сравнивала модели, описывающие кругооборот биогенных ресурсов (азота или азота и фосфора) в замкнутой системе, включающей одну группу фитопланктона, питательные элементы и детрит. При использовании клеточной квоты удельная скорость роста задавалась формулой Друпа, при этом в случае двух ресурсов использовался принцип минимума Либиха. Оказалось, что модели с использованием клеточной квоты дают большее число возможных сценариев развития фитопланктона и позволяют описывать ситуации, реально встречающиеся в природе, например, ситуацию нарастания биомассы фитопланктона в условиях почти нулевой концентрации биогенного элемента в среде или ситуацию резкого скачка отношения N:P в среде в момент кульминации цветения.
В настоящее время среди исследователей установилось мнение, что модель Моно адекватна при описании устойчивого роста водорослей в стационарных условиях, подобных хемостату, а для описания процесса роста при изменяющихся внешних условиях, например, при моделировании внутригодовой сукцессии фитопланктона, необходима более сложная модель, выбранная в соответствии с предположением зависимости скорости роста от клеточных концентраций ресурсов. Одной из ситуаций, где при моделировании необходимо привлечение концепции клеточной квоты, является накопительное культивирование.
3.1.2. Дифференциальные уравнения в микробиологии. Дифференциальные уравнения, описывающие и концентрации микробной биомассы (x) и концентрации лимитирующего субстрата (s) в условиях хемостатного культивирования, которые были выведены Ж.Моно исходя из условий материального баланса, составили первую модель роста микробных популяций (Паников, 1991)
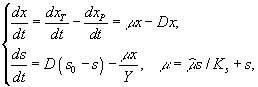
где xP – концентрация микроорганизмов в приемнике, куда сливается нарастающая бактериальная суспензия; xT – суммарная концентрация микроорганизмов (xT = x + xP); μ – удельная скорость роста, ![]() – максимальная скорость роста; s – концентрация лимитирующего ресурса в среде; Ks – константа полунасыщения при лимитировании данным субстратом; s0 – величина концентрации лимитирующего субстрата на входе в культиватор; D – скорость разбавления, равная отношению скорости поступления питательной среды к объему культуры; Y – экономический коэффициент (выход биомассы на единицу потребленного субстрата).
– максимальная скорость роста; s – концентрация лимитирующего ресурса в среде; Ks – константа полунасыщения при лимитировании данным субстратом; s0 – величина концентрации лимитирующего субстрата на входе в культиватор; D – скорость разбавления, равная отношению скорости поступления питательной среды к объему культуры; Y – экономический коэффициент (выход биомассы на единицу потребленного субстрата).
Принципиальной особенностью данной открытой системы является возможность установления динамического равновесия. В установившемся состоянии μ = D, а ![]() (
(![]() – постоянные значения стационарного состояния).
– постоянные значения стационарного состояния).
Необходимо также отметить, что при превышении скорости разбавления D критического значения ![]() наступает режим вымывания, т.е. скорость потока настолько велика, что прирост биомассы не может компенсировать ее отток. Нулевое значение концентрации биомассы и равенство концентрации лимитирующего ресурса в среде значению последней на входе в культиватор является неустойчивым положением равновесия при скорости разбавления ниже критической и устойчивым при больших скоростях
наступает режим вымывания, т.е. скорость потока настолько велика, что прирост биомассы не может компенсировать ее отток. Нулевое значение концентрации биомассы и равенство концентрации лимитирующего ресурса в среде значению последней на входе в культиватор является неустойчивым положением равновесия при скорости разбавления ниже критической и устойчивым при больших скоростях ![]() (Романовский и др., 1971).
(Романовский и др., 1971).
Для различных методов проточного культивирования динамика концентрации биомассы микроорганизмов описывается соответствующими дифференциальными уравнениями (Паников, 1991).
1) Турбидостат 
![]() – оптическая плотность.
– оптическая плотность.
2) pH-стат 
3) Хемостат с возвратом 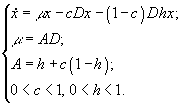
4) Батарея хемостатов 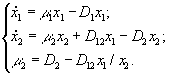
Наряду с различными способами гомогенного непрерывного культивирования для лабораторных исследований применяют и другие методы:
1) Непрерывное культивирование без выноса микробных клеток:
а) периодический диализ 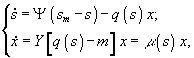
б) непрерывный диализ 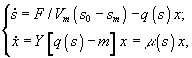
в) периодическая культура с подпиткой (ПКП)
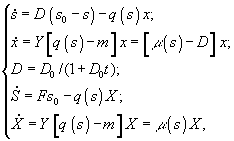
г) периодическая культура с газовым питанием
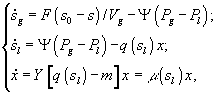
![]() – величины концентрации лимитирующего субстрата в диализном сосуде, в газовой и жидкой фазах; Pg и Pl – парциальные давления; Ψ – коэффициент массопереноса;
– величины концентрации лимитирующего субстрата в диализном сосуде, в газовой и жидкой фазах; Pg и Pl – парциальные давления; Ψ – коэффициент массопереноса; ![]() – объемы диализного сосуда, воздушной фазы ферментера,; S, X – количество субстрата и биомассы в расчете на всю культуру (s, x – в расчете на единицу объема); F – поток лимитирующего субстрата; s0 – исходная концентрация лимитирующего субстрата; q(s) - скорость потребления лимитирующего субстрата.
– объемы диализного сосуда, воздушной фазы ферментера,; S, X – количество субстрата и биомассы в расчете на всю культуру (s, x – в расчете на единицу объема); F – поток лимитирующего субстрата; s0 – исходная концентрация лимитирующего субстрата; q(s) - скорость потребления лимитирующего субстрата.
2) Непрерывная гомогенная культура с “залповой” подачей лимитирующего субстрата, при которой в обычную хемостатную культуру непрерывно поступает питательная среда, но без одного из существенных компонентов, который вносится в виде малой порции концентрированного раствора A через регулярные достаточно большие промежутки времени ![]() . В этом случае динамика лимитирующего субстрата s и микробной биомассы x описывается системой
. В этом случае динамика лимитирующего субстрата s и микробной биомассы x описывается системой
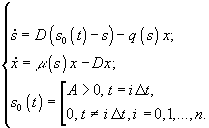
Применяют также обычное гомогенное периодическое культивирование и воспроизведение пространственно-неоднородного роста микроорганизмов:
а) культура полного вытеснения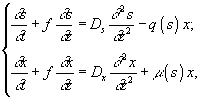
f – скорость перемещения малого элемента культуральной жидкости по пространственной координате z; Ds и Dx – коэффициенты диффузии для субстрата и клеток;
б) проточный реактор с иммобилизованными клетками

Одной из областей применения дифференциальных уравнений в моделировании микробиологических сообществ является анализ возможных стационарных состояний смешанной культуры. Примером такого исследования может служить работа И.Р.Спивака и Дж.С.Рокема (Spivak, Rokem, 1995), в которой изучалось взаимодействие двух видов гетеротрофных бактерий, конкурирующих за промежуточные внеклеточные метаболиты, выделяемые бактериями-метилотрофами. Эти метаболиты, в свою очередь, являются ингибиторами для метилотрофных бактерий. Система уравнений, описывающих такое сообщество, включает в себя выражения для динамики бактериальных биомасс, а также динамики концентрации кислорода, метанола, этанола, ацетата в ферментационной жидкой среде

Удельная скорость роста видов задается формулами

Скорость потребления субстрата cij может быть представлена как 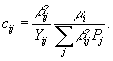 В вышеприведенных уравнениях использованы обозначения: xi (i = 1,2,3) – бактериальная биомасса метилотрофа и двух видов гетеротрофов;
В вышеприведенных уравнениях использованы обозначения: xi (i = 1,2,3) – бактериальная биомасса метилотрофа и двух видов гетеротрофов; ![]() – удельная и максимальная удельная скорости роста i-го вида; D – скорость вымывания; s1, Pj (j = 1…4) – концентрации кислорода, СО2, метанола, этанола и ацетата в среде соответственно (верхний индекс * обозначает концентрацию вещества в стацонарном состоянии); KLi (i=1,2) – коэффициент переноса массы для кислорода и СО2 соответственно; Yij (i = 1,2,3; j = 1,3,4) – выход бактерий вида i на j-м субстрате;
– удельная и максимальная удельная скорости роста i-го вида; D – скорость вымывания; s1, Pj (j = 1…4) – концентрации кислорода, СО2, метанола, этанола и ацетата в среде соответственно (верхний индекс * обозначает концентрацию вещества в стацонарном состоянии); KLi (i=1,2) – коэффициент переноса массы для кислорода и СО2 соответственно; Yij (i = 1,2,3; j = 1,3,4) – выход бактерий вида i на j-м субстрате; ![]() (i = 1,2,3; j = 1,3,4) – коэффициент подобия бактерий вида i субстрату j;
(i = 1,2,3; j = 1,3,4) – коэффициент подобия бактерий вида i субстрату j; ![]() (i = 1,2,3; j = 1,…,4) – коэффициент чувствительности бактерий вида i к метабоиту n;
(i = 1,2,3; j = 1,…,4) – коэффициент чувствительности бактерий вида i к метабоиту n; ![]() – удельная скорость выделения внеклеточного метаболита j бактериями вида i.
– удельная скорость выделения внеклеточного метаболита j бактериями вида i.
В работе подробно изучены 6 различных способов лимитирования: метилотрофные бактерии во всех случаях лимитируются конценрацией растворенного кислорода, два вида гетеротрофов лимитируются либо оба промежуточными внеклеточными метаболитами, либо оба максимальной удельной скоростью роста, либо оба конценрацией растворенного кислорода, либо сочетнием двух разных лимитирующх факторов. Среди полученных результатов необходимо отметить возможность существования устойчивой культуры, состоящей из трех бактериальных видов, конкурирующих за кислород.
Важным направлением микробиологических исследований является изучение генетической неоднородности клеток в непрерывной культуре (Паников, 1991). Хемостатная модель, описывающая ситуацию, когда исходная форма дает целый веер мутаций с реверсиями, записывается следующим образом

здесь xi – концентрация i-го варианта; x – суммарная концентрация клеток в хемостатной культуре; ![]() – матрица всех возможных переходов между вариантами xi.
– матрица всех возможных переходов между вариантами xi.
Попытка провести количественную оценку возможного вклада процесса диссоциации – расщепления однородной популяции бактерий на варианты, различающиеся генетическими, физиолого-биологическими и морфологическими свойствами – в формирование гетерогенной биологической популяции была предпринята в работах Е.С.Милько с соавторами (Милько, Егоров, 1991; Drozdov-Tikhomirov at al., 1989). Математическая модель для таких процессов выражается системой уравнений
 где
где ![]() – биомассы R-, S-, и M-диссоциантов соответственно; s – концентрация субстрата в среде; Ki – константа полунасыщения вида i субстратом s; D – скорость протока; s0 – начальная концентрация субстрата; Y – экономический коэффициент, принятый в модели одинаковым для всех диссоциантов;
– биомассы R-, S-, и M-диссоциантов соответственно; s – концентрация субстрата в среде; Ki – константа полунасыщения вида i субстратом s; D – скорость протока; s0 – начальная концентрация субстрата; Y – экономический коэффициент, принятый в модели одинаковым для всех диссоциантов; ![]() – частота диссоциативных переходов.
– частота диссоциативных переходов.
Важным фактором баланса роста клеток являются затраты энергии на их поддержание. Эти затраты складываются из расходов на повторный синтез нестабильных макромолекул и клеточных структур, на поддержание градиентов веществ между клеткой и окружающей средой, а также между компартментами клетки, на подвижность клеток (если они ею обладают), процесс сброса избыточной энергии, поступающей в клетки (Минкевич и др., 1998). Удельная скорость затрат энергии на поддержание ![]() зависит от вида микроорганизмов, субстрата, температуры, возможно, и от других факторов. Влияние концентраций ингибирующих субстратов этанола и цинка на величину
зависит от вида микроорганизмов, субстрата, температуры, возможно, и от других факторов. Влияние концентраций ингибирующих субстратов этанола и цинка на величину ![]() клеток этанолассимилирующих дрожжей Candida valida изучалось в работе И.Г.Минкевича с соавторами (Минкевич и др., 1998). В результате исследования были получены формулы для скорости потребления энергетического субстрата qS и удельной скорости роста
клеток этанолассимилирующих дрожжей Candida valida изучалось в работе И.Г.Минкевича с соавторами (Минкевич и др., 1998). В результате исследования были получены формулы для скорости потребления энергетического субстрата qS и удельной скорости роста ![]() , которые отражают качественные особенности влияния данных субстратов на рост культуры – лимитирования, существования наклонного плато и субстратного ингибирования:
, которые отражают качественные особенности влияния данных субстратов на рост культуры – лимитирования, существования наклонного плато и субстратного ингибирования:

Член ![]() , описывающий влияние этанола, имеет вид
, описывающий влияние этанола, имеет вид
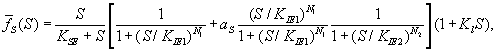
член ![]() , описывающий влияние цинка на скорость потребления энергетического субстрата qS , записывают как
, описывающий влияние цинка на скорость потребления энергетического субстрата qS , записывают как
![]()
Влияние концентрации цинка в окружении клеток на удельную скорость затрат энергии на ![]() описывают выражениями
описывают выражениями
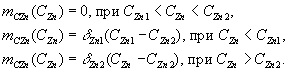
В приведенных формулах CZn обозначает внешнюю концентрацию цинка, CZn1 и CZn2 – внутреннюю концентрацию цинка, S – концентрацию этанола; ![]() – максимальный выход роста; b – константа;
– максимальный выход роста; b – константа; ![]()
![]() – физиологические параметры культуры Candida valida.
– физиологические параметры культуры Candida valida.
Предлагаемая модель обеспечивает высокую степень соответствия экспериментальным данным и объясняет многие особенности поведения культуры. По мнению авторов, описанные зависимости могут применяться ко многим органическим и минеральным субстратам, в том числе обладающим токсическим действием на микроорганизмы.
3.2. Концепция лимитирующих факторов. Существование и успех любого организма или любой группы организмов зависит от комплекса определенных условий. Любое условие, приближающееся к пределу толерантности или превышающее его, называется лимитирующим условием, или лимитирующим фактором (Одум, 1986а). Исследование влияния разнообразных факторов на рост растений позволило Ю.Либиху сделать вывод о том, что “рост растений зависит от того элемента питания, который присутствует в минимальном количестве” (Liebig, 1840). В последствии этот результат получил название “принципа минимума Либиха”. Дальнейшие исследования показали, что для успешного применения этого принципа необходимо учитывать взаимодействие факторов (например, высокая концентрация, доступность или воздействие не лимитирующего фактора может изменить скорость потребления элемента питания, содержащегося в минимальном количестве), а также ограничение, согласно которому, закон Либиха строго применим только в условиях стационарного состояния, т.е. когда приток и отток энергии и вещества сбалансированы (Одум, 1986а).
При практическом использовании принципа Либиха важнейшим моментом является выявление лимитирующего элемента питания. Сам Ю.Либих определял лимитирующий ресурс по соотношению между запасами веществ в организме и в окружающей среде (Алексеев и др., 1992).
Использование в математических моделях зависимости скорости роста популяции как функции одного элемента питания μ(s) допустимо только для систем с установившимся характером лимитирования. Однако, смена факторов лимитирования встречается повсеместно и представляет как теоретический, так и практический интерес. Существует несколько различных подходов к теоретическому описанию механизма смены факторов, лимитирующих рост популяции. Наиболее распространенными являются модели, в которых предполагается, что в любой момент времени рост контролируется только одним элементом питания. Среди таких моделей можно выделить два варианта определения момента смены лимитирования (Алексеев и др., 1992).
В первом варианте лимитирующим считается элемент питания, дающий наименьшую скорость роста популяции. Например, для модели Моно смена лимитирующего элемента питания с s1 на s2 записывается следующим образом
![]() если
если ![]()
![]() если
если ![]()
причем смена лимитирования может происходить только при неравенстве максимальных скоростей роста: ![]() .
.
Во втором варианте лимитирующим является элемент питания, для которого отношение “концентрация в среде/концентрация в организме” оказывается наименьшим среди подобных отношений для других незаменимых компонентов питания. Смена лимитирования в этом варианте записывается следующим образом
![]() если
если ![]()
![]() если
если ![]()
где ![]() и
и ![]() – удельное содержание первого и второго элементов в организмах популяции соответственно.
– удельное содержание первого и второго элементов в организмах популяции соответственно.
Для обеих моделей на плоскости ![]() (
(![]() – концентрация j-го питательного вещества на входе системы) выделяют три области стационарных состояний с определенным типами лимитирования в зависимости от запасов биогенных элементов
– концентрация j-го питательного вещества на входе системы) выделяют три области стационарных состояний с определенным типами лимитирования в зависимости от запасов биогенных элементов ![]() и
и ![]() : область вымирания популяции; область, где в стационарном состоянии рост популяции лимитирует элемент s1, и область лимитирования элементом s2. Однако форма границ областей в моделях заметно различаются (Алексеев и др., 1992). Экспериментальная проверка реалистичности двух механизмов смены лимитирования (лимитирование культуры микроводоросли Chlorella sp. фосфатами и нитратами) показала (Алексеев, Мурзин, 1982), что более корректно механизм жесткой смены лимитирования описывает вторая модель, которая совпадает с представлениями Либиха.
: область вымирания популяции; область, где в стационарном состоянии рост популяции лимитирует элемент s1, и область лимитирования элементом s2. Однако форма границ областей в моделях заметно различаются (Алексеев и др., 1992). Экспериментальная проверка реалистичности двух механизмов смены лимитирования (лимитирование культуры микроводоросли Chlorella sp. фосфатами и нитратами) показала (Алексеев, Мурзин, 1982), что более корректно механизм жесткой смены лимитирования описывает вторая модель, которая совпадает с представлениями Либиха.
3.2.1. Применение различных способов формализации закона минимума Либиха в моделях с дифференциальными уравнениями. Лимитирующим считается элемент питания, дающий наименьшую скорость роста популяции. Этот подход к определению момента переключения лимитирующего фактора применялся в модели конкуренции двух видов организмов за два взаимонезаменяемых ресурса в режиме хемостатного культивирования (Абросов, Ковров, 1977). Динамика биомасс видов x1, x2 и питательных веществ s1, s2, описывается уравнениями

где xi – концентрация биомассы i-го вида; D – скорость протока в хемостате; ![]() ,
, ![]() – удельная скорость роста организмов;
– удельная скорость роста организмов; ![]() – концентрация j-го вещества в поступающей в реактор питательной среде; sj – концентрация элементов питания в реакторе; Yij – экономический коэффициент использования j-го вещества организмами i-го вида;
– концентрация j-го вещества в поступающей в реактор питательной среде; sj – концентрация элементов питания в реакторе; Yij – экономический коэффициент использования j-го вещества организмами i-го вида; ![]() – коэффициент приспособленности i-го вида к j-му ресурсу. В уравнениях используется кусочно-линейная аппроксимация формулы Моно.
– коэффициент приспособленности i-го вида к j-му ресурсу. В уравнениях используется кусочно-линейная аппроксимация формулы Моно.
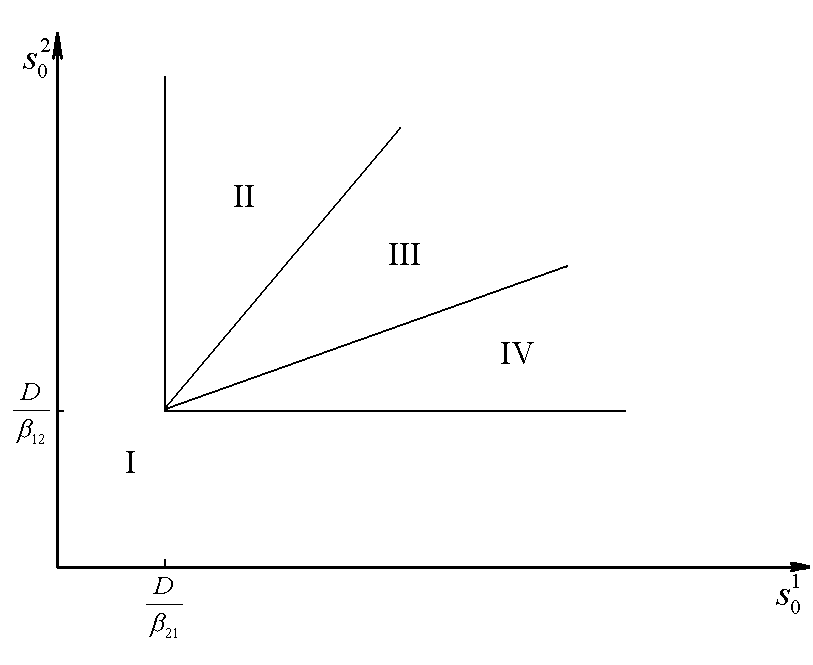
Рис.1. Первый вид лимитируется ресурсом s1, второй – ресурсом s2. I – область вымирания обоих видов, II – область существования первого вида, III – область сосуществования, IV – область существования второго вида. D – скорость протока в хемостате;![]() – коэффициент приспособленности i-го вида к j-му ресурсу;
– коэффициент приспособленности i-го вида к j-му ресурсу; ![]() – концентрация j-го вещества в поступающей в реактор питательной среде
– концентрация j-го вещества в поступающей в реактор питательной среде
Исследование стационарных состояний системы дало следующие результаты. Если оба вида потребляют преимущественно один и тот же ресурс, то в системе становится доминирующим один вид, тот, у которого максимален коэффициент β по лимитирующему ресурсу. Если же виды потребляют преимущественно разные ресурсы, то результат конкурентной борьбы зависит от значений ![]() – концентраций вещества j в поступающей в реактор среде. Пусть, для определенности
– концентраций вещества j в поступающей в реактор среде. Пусть, для определенности ![]() ,
, ![]() . Тогда в стационарном состоянии скорости роста популяций определяется формулами
. Тогда в стационарном состоянии скорости роста популяций определяется формулами ![]() ;
; ![]() . Отсюда следует, что на плоскости
. Отсюда следует, что на плоскости ![]() можно выделить область вымывания обоих видов
можно выделить область вымывания обоих видов 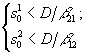 (для значений концентраций
(для значений концентраций ![]() из этой области
из этой области ![]() ). Для остальных значений концентраций вещества
). Для остальных значений концентраций вещества ![]() возможны три варианта устойчивого равновесия: сосуществование обоих видов, доминирование первого вида, доминирование второго вида (рис.1). Границы области сосуществования задаются прямыми
возможны три варианта устойчивого равновесия: сосуществование обоих видов, доминирование первого вида, доминирование второго вида (рис.1). Границы области сосуществования задаются прямыми  и
и 
Лимитирующим является элемент питания, для которого отношение “концентрация в среде/концентрация в организме” – наименьшее по сравнению с другими незаменимыми компонентами питания. Этот механизм лимитирования применяли в модели конкуренции между микроводорослями в замкнутой системе (Алексеев, Сазыкина, 1981; Алексеев и др., 1992). Предполагали, что элементный химический состав организмов фиксирован; каждый вид потребляет биогенные вещества в пропорциях, соответствующих соотношению элементов в клетках водоросли. Динамика биомасс в случае конкуренции двух видов за два незаменимых ресурса s1 и s2 описывается уравнениями
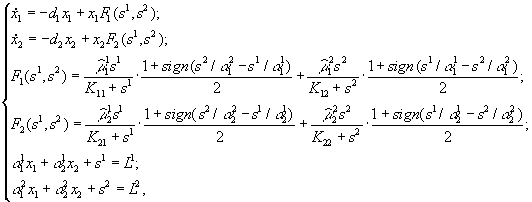
где xi – плотность биомассы i-го вида; ![]() – содержание k-го элемента в клетках i-го вида; di – коэффициенты естественной смертности;
– содержание k-го элемента в клетках i-го вида; di – коэффициенты естественной смертности; ![]() – максимальные скорости прироста биомассы вида i при лимитровании веществом j;
– максимальные скорости прироста биомассы вида i при лимитровании веществом j; ![]() – константы полунасыщения вида i при лимитровании веществом j; Lj – общие запасы первого и второго элементов питания в системе. Функция sign(x) принимает значение, равное 1 при x > 0, равное – 1 при x < 0 и равное 0 при x = 0.
– константы полунасыщения вида i при лимитровании веществом j; Lj – общие запасы первого и второго элементов питания в системе. Функция sign(x) принимает значение, равное 1 при x > 0, равное – 1 при x < 0 и равное 0 при x = 0.
Области устойчивости зависят от Lj, x1(0), x2(0). Пусть ![]() . В зависимости от того, какой ресурс является лимитирующим, положительный квадрат плоскости общих запасов биогенов
. В зависимости от того, какой ресурс является лимитирующим, положительный квадрат плоскости общих запасов биогенов ![]() разбивается на три сектора (рис.2).
разбивается на три сектора (рис.2).
 Рис.2. Разбиение плоскости общих запасов биогенных элементов в зависимости от того, какое вещество является лимитирующим для каждого из видов.
Рис.2. Разбиение плоскости общих запасов биогенных элементов в зависимости от того, какое вещество является лимитирующим для каждого из видов. ![]() – содержание k-го элемента в клетках i-го вида; L1, L2 – общие запасы питательных веществ. В области I лимитирующим является вещество s1, в области III – вещество s2, в II – возможны четыре комбинации: популяции лимитируются разными или одинаковыми ресурсами
– содержание k-го элемента в клетках i-го вида; L1, L2 – общие запасы питательных веществ. В области I лимитирующим является вещество s1, в области III – вещество s2, в II – возможны четыре комбинации: популяции лимитируются разными или одинаковыми ресурсами
В первом – лимитирующим является вещество s1, в третьем – вещество s2, во втором – возможны четыре комбинации: популяции лимитируются разными или одинаковыми биогенами. Границы секторов задаются осями координат и лучами ![]() . В зависимости от величин минимальных концентраций
. В зависимости от величин минимальных концентраций ![]() ресурсов s1 и s2 в среде, необходимых для существования соответственно видов x1 и x2 сектора I и III разбиваются прямыми L1= min(
ресурсов s1 и s2 в среде, необходимых для существования соответственно видов x1 и x2 сектора I и III разбиваются прямыми L1= min(![]() ,
,![]() ) и L2 = min (
) и L2 = min (![]() ,
, ![]() ) на зону доминирования популяции, наиболее неприхотливой по отношению к биогену s1 и s2, соответственно, (рис. 3 а, б, в, г)
) на зону доминирования популяции, наиболее неприхотливой по отношению к биогену s1 и s2, соответственно, (рис. 3 а, б, в, г)
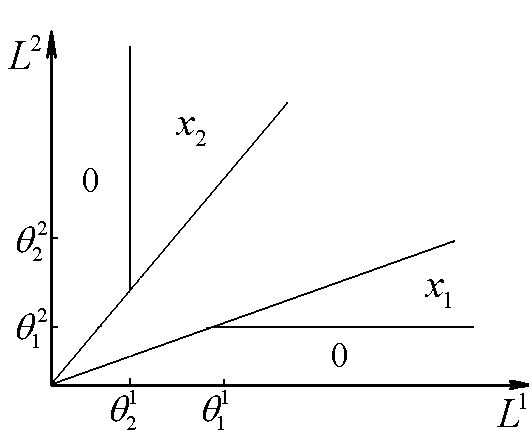
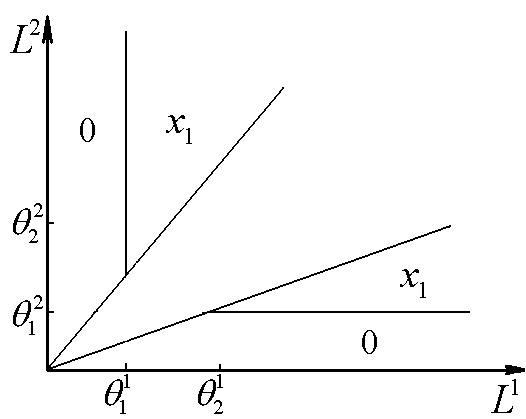 (
(![]() ,
,![]() ,
,![]() ,
,![]() ).
).
а) б)
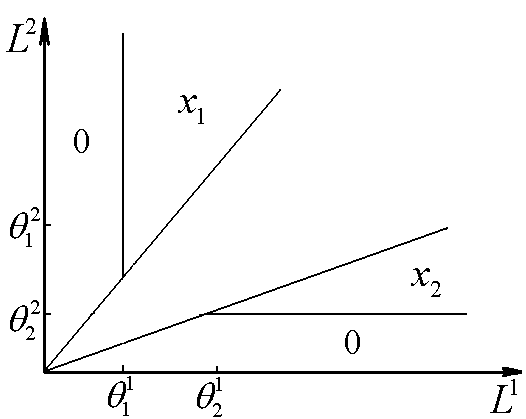
в) г)
Рис.3. В области 0 происходит вымирание обоих видов, в области x1 доминирует первый вид, в области x2 – второй. ![]() – минимальные концентрации биогенов s1 и s2 в среде, необходимые для существования соответственно видов x1 и x2. L1, L2 – общие запасы питательных веществ
– минимальные концентрации биогенов s1 и s2 в среде, необходимые для существования соответственно видов x1 и x2. L1, L2 – общие запасы питательных веществ
 Далее следует изучение сектора II. Случай, когда вид x1 лимитируется ресурсом s2, а вид x2 – ресурсом s1, не обладает устойчивым состоянием равновесия. Вариант динамики, когда оба вида ограничены веществом s2, имеет одно стационарное состояние – доминирование популяции вида x2. На плоскости (L1, L2) эта ситуация реализуется в секторе с вершиной в точке (
Далее следует изучение сектора II. Случай, когда вид x1 лимитируется ресурсом s2, а вид x2 – ресурсом s1, не обладает устойчивым состоянием равновесия. Вариант динамики, когда оба вида ограничены веществом s2, имеет одно стационарное состояние – доминирование популяции вида x2. На плоскости (L1, L2) эта ситуация реализуется в секторе с вершиной в точке (![]() ), у которого нижний луч совпадает с нижней границей сектора II, а верхний луч – параллелен верхней границе. Состояние устойчиво, если
), у которого нижний луч совпадает с нижней границей сектора II, а верхний луч – параллелен верхней границе. Состояние устойчиво, если ![]() (рис. 4).
(рис. 4).
Рис.4. В области x2 доминирует второй вид. ![]() – минимальные концентрации биогенов s1 и s2 в среде, необходимые для существования соответственно видов x1 и x2. L1, L2 – общие запасы питательных веществ
– минимальные концентрации биогенов s1 и s2 в среде, необходимые для существования соответственно видов x1 и x2. L1, L2 – общие запасы питательных веществ
Если оба вида лимитируются биогеном s1, равновесным является доминирование вида x1. Область доминирования изображается сектором с вершиной в точке (![]() ), верхним лучем, совпадающим с верхней границей сектора II, нижним – параллельным нижней границе. Состояние устойчиво, если
), верхним лучем, совпадающим с верхней границей сектора II, нижним – параллельным нижней границе. Состояние устойчиво, если ![]() (рис. 5).
(рис. 5).
При лимитировании популяции вида x1 биогеном s2, а популяции вида x2 – биогеном s1 конкуренция может иметь четыре исхода: вымирание обоих видов, сосуществование, доминирование одного или другого. Состояние вымирания устойчиво при![]() ,
, ![]() .
.
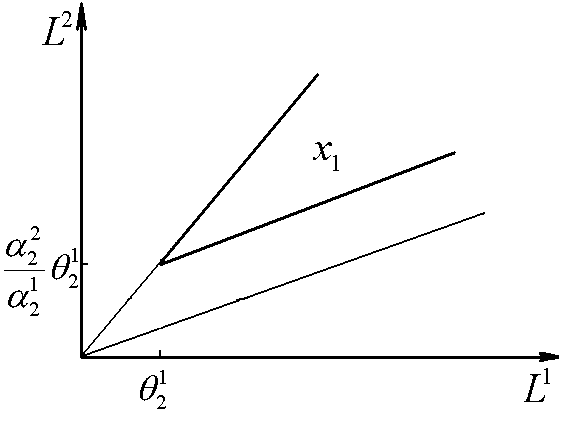 Рис.5. В области x1 доминирует первый вид.
Рис.5. В области x1 доминирует первый вид. ![]() – минимальные концентрации биогенов s1 и s2 в среде, необходимые для существования соответственно видов x1 и x2. L1, L2 – общие запасы питательных веществ
– минимальные концентрации биогенов s1 и s2 в среде, необходимые для существования соответственно видов x1 и x2. L1, L2 – общие запасы питательных веществ
Область доминирования вида x2 пуста, если точка (![]() ) принадлежит сектору I, и имеет вид полосы, примыкающей к верхнему лучу среднего сектора, в остальных случаях. Исход устойчив, если верны неравенства
) принадлежит сектору I, и имеет вид полосы, примыкающей к верхнему лучу среднего сектора, в остальных случаях. Исход устойчив, если верны неравенства
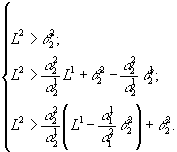
Область доминирования вида x1 пуста, если точка (![]() ) лежит в III секторе, и ограничена полосой, примыкающей к нижней границе среднего сектора, в других случаях. Условия положительности и устойчивости
) лежит в III секторе, и ограничена полосой, примыкающей к нижней границе среднего сектора, в других случаях. Условия положительности и устойчивости
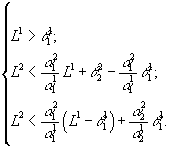
Ситуация сосуществования видов возможна только, когда точка (![]() ) лежит в секторе II. На плоскости
) лежит в секторе II. На плоскости ![]() область значений L1 и L2, при которых виды могут сосуществовать, задается сектором с вершиной в точке (
область значений L1 и L2, при которых виды могут сосуществовать, задается сектором с вершиной в точке (![]() ) и сторонами, параллельными сторонам сектора II (рис. 6).
) и сторонами, параллельными сторонам сектора II (рис. 6).
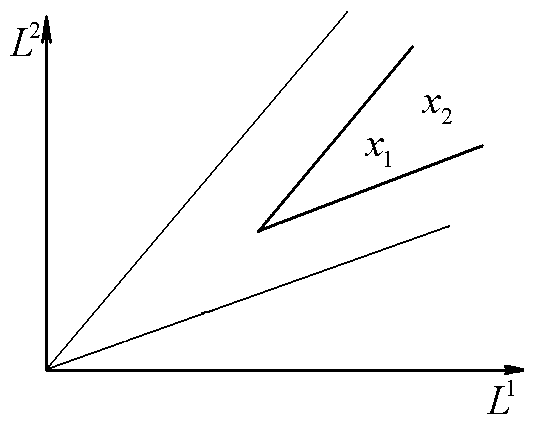 Рис.6. В области x1, x2 сосуществуют два вида. L1, L2 – общие запасы питательных веществ
Рис.6. В области x1, x2 сосуществуют два вида. L1, L2 – общие запасы питательных веществ
Таким образом, можно выделить 12 различных способов заполнения плоскости биогенов (три варианта принадлежности точки (![]() ) одному из секторов, каждый из которых делится на четыре по положению значений
) одному из секторов, каждый из которых делится на четыре по положению значений ![]() на оси L1 и
на оси L1 и ![]() на оси L2). Причем, кроме случаев, когда области доминирования прилегают друг к другу, существуют варианты наложений различных исходов (в этой ситуации результат зависит от начального значения биомасс), а также разрывы, внутри которых происходят колебания.
на оси L2). Причем, кроме случаев, когда области доминирования прилегают друг к другу, существуют варианты наложений различных исходов (в этой ситуации результат зависит от начального значения биомасс), а также разрывы, внутри которых происходят колебания.
3.2.2. Индикатор лимитирующего ресурса. Модель динамики фитопланктона, включающая в себя закон минимума Либиха, была проанализирована в работе С.Леговича и А.Крузадо (Legovic, Cruzado, 1997). Результатом исследования стал индикатор для определения лимитирующего ресурса. Модель описывается системой из 2n + 1 линейных дифференциальных уравнений
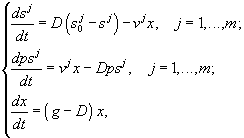
где 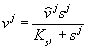 (зависимость Михаэлиса-Ментена-Моно скорости поглощения
(зависимость Михаэлиса-Ментена-Моно скорости поглощения ![]() от концентрации элемента питания sj),
от концентрации элемента питания sj), ![]() ,
,![]() ; m – количество ресурсов, sj – концентрация j-го ресурса в среде, psj – концентрация j-го ресурса в фитопланктоне, x – плотность клеток, D – скорость промывания реактора,
; m – количество ресурсов, sj – концентрация j-го ресурса в среде, psj – концентрация j-го ресурса в фитопланктоне, x – плотность клеток, D – скорость промывания реактора, ![]() – концентрация i-го ресурса во входящем потоке,
– концентрация i-го ресурса во входящем потоке, ![]() и
и ![]() – максимальная скорость поглощения и константа полунасыщения фитопланктона при потреблении j-го ресурса,
– максимальная скорость поглощения и константа полунасыщения фитопланктона при потреблении j-го ресурса, ![]() – максимальная скорость деления,
– максимальная скорость деления, ![]() – квота j-го ресурса в живых, но неделящихся клетках (все параметры – положительные константы).
– квота j-го ресурса в живых, но неделящихся клетках (все параметры – положительные константы).
Считается, что потребление ресурсов независимо друг от друга, клетки фитопланктона содержат, по крайней мере, m различных типов “мест для хранения питательного вещества”.
Система уравнений определяет следующие состояния равновесия: тривиальное ![]() = 0,
= 0, ![]() = 0, x* = 0, j= 1,...,m, которое не существует в данной модели; состояние
= 0, x* = 0, j= 1,...,m, которое не существует в данной модели; состояние ![]() =
= ![]() ,
, ![]() = 0, x* = 0 – существует, но не устойчиво при определенных значениях параметра; нетривиальное состояние равновесия определяется из условия
= 0, x* = 0 – существует, но не устойчиво при определенных значениях параметра; нетривиальное состояние равновесия определяется из условия ![]() , j = 1,...m и
, j = 1,...m и ![]() > 0,
> 0, ![]() > 0, x* > 0,
> 0, x* > 0, ![]() >
> ![]() . Данные условия порождают систему уравнений для нахождения состояния равновесия
. Данные условия порождают систему уравнений для нахождения состояния равновесия
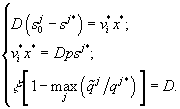
Предполагая, что 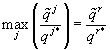 , вычисляются величины
, вычисляются величины ![]() , а затем – величины
, а затем – величины ![]()
Однако, в таком решении нет пути, по которому можно определить, какой ресурс является лимитирующим. При проведении эксперимента этот вопрос легко решается, а для аналитического решения авторы предлагают следующий метод: найти все потенциальные состояния равновесия, предполагая каждый ресурс лимитирующим, а затем выбрать тот, для которого значение плотности клеток в стационарном состоянии x* – минимальное, причем, если рост фитопланктона ограничивают несколько ресурсов, то стационарные плотности x* для этих веществ будут иметь одинаковые, наименьшие по сравнению с не лимитирующими, значения.
Предложенный метод позволяет ввести индикатор, для определения лимитирующего ресурса
![]() где
где ![]() .
.
3.2.3. Развитие концепции лимитирующих факторов. В реальной системе скорость первичной ассимиляции на всех этапах продукционного цикла может контролироваться периодически сменяющимися элементами, лимитирующими фотосинтез, а также находиться под влиянием совокупного действия факторов. Для преодоления связанных с этим трудностей Ю.А.Домбровский с коллегами (1990) предлагает следующее развитие концепции лимитирования. Во-первых, проводится распространение понятия лимитирования на первичную продукцию или ассимиляцию: для оценки лимитирующего элемента ![]() (в предположении независимого действия биогенов) используется выражение
(в предположении независимого действия биогенов) используется выражение
![]() ,
, ![]()
(функция arg принимает значение того индекса, при котором реализуется минимум выражения) здесь А – годовая ассимиляция в единицах органического углерода; bj – весовое отношение, в котором углерод соединяется с другими биогенными элементами в ходе реакции фотосинтеза; cj – скорость первичного круговорота при лимитировании j-м элементом; sj – количество j-го вещества, участвующего в первичном круговороте.
Во-вторых, предлагается отказаться от гипотезы независимого действия факторов и включить закон Митчерлиха наряду с законом минимума и пределами толерантности в единую концептуальную схему лимитирования. Пусть элементарный поток вещества pj описывается общим выражением ![]() . Лимитирующий фактор j0 определяется из условия
. Лимитирующий фактор j0 определяется из условия
![]() (3.4)
(3.4)
(функция arg принимает значение того индекса, при котором реализуется максимум выражения) которое означает, что лимитирующий фактор вызывает наибольшее изменение потока pj на единицу своего относительного изменения.
Применение критерия (3.4) к годовой первичной ассимиляции, играющей роль интегральной функции отклика всей водной экосистемы, дает критерий выбора лимитирующего элемента для модели первичного круговорота
![]()
![]()
3.3. Моделирование конкуренции за ресурсы. Одним из факторов, влияющих на формирование структуры экологических сообществ, является конкуренция. Конкуренция в самом широком смысле – это взаимодействие организмов, стремящихся получить один и тот же ресурс.
Конкурентное взаимодействие может касаться пространства, пищи или биогенных элементов, света, зависимости от хищников и т.д. Межвидовая конкуренция может привести либо к установлению равновесия между двумя видами, либо к замене популяции одного вида на популяцию другого, либо к тому, что один вид вытеснит другой в иное место или же заставит его перейти на использование иной пищи. При конкуренции близкородственных или сходных в иных отношениях видов наблюдается тенденция к их экологическому разделению (близкородственные, ведущие сходный образ жизни и обладающие сходной морфологией организмы, обитают в разных местах или используют разные ресурсы или разное время активности при занятии одного местообитания). Эта тенденция получила известность как принцип конкурентного исключения (принцип Гаузе) (Одум, 1986б).
3.3.1. Совместное потребление ресурсов питания организмами одного трофического уровня. Случай конкуренции за взаимонезаменяемые ресурсы характерен для автотрофного трофического уровня, составленного, например, одноклеточными водорослями. В работе Н.С.Абросова с коллегами (Абросов, Ковров, 1977) подробно исследована модель динамики концентрации клеток
 (3.5)
(3.5)
(Обозначения те же, что в формулах п. 3.1.1 – (3.1) и др.)
Стационарное состояние системы (3.5) определяется уравнениями
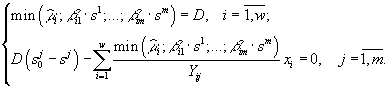
Исследование модели в случае конкуренции двух видов за два ресурса позволяет (Абросов, 1999) построить разбиение пространства начальных концентраций питательных веществ ![]() на области, в которых реализуются различные режимы лимитирования и исходы конкуренции (рис.7). В области I реализуется преимущество первого вида при лимитировании обоих конкурентов субстратом s1; в области Ia побеждает первый вид при лимитировании первого вида субстратом s2, а второго – субстратом s1; в области II конкуренцию выигрывает второй вид при лимитировании обоих конкурентов субстратом s2; в области IIa выигрывает второй вид при лимитировании второго вида субстратом s1, а первого вида – субстратом s2. В области III реализуется устойчивое сосуществование конкурентов при лимитировании первого вида субстратом s2, а второго вида – субстратом s1. Область IV, границей которой является ломаная линия MNAPQ, – область нулевой биомассы сообщества. Коэффициенты угла наклона прямых, являющихся границами областей, есть
на области, в которых реализуются различные режимы лимитирования и исходы конкуренции (рис.7). В области I реализуется преимущество первого вида при лимитировании обоих конкурентов субстратом s1; в области Ia побеждает первый вид при лимитировании первого вида субстратом s2, а второго – субстратом s1; в области II конкуренцию выигрывает второй вид при лимитировании обоих конкурентов субстратом s2; в области IIa выигрывает второй вид при лимитировании второго вида субстратом s1, а первого вида – субстратом s2. В области III реализуется устойчивое сосуществование конкурентов при лимитировании первого вида субстратом s2, а второго вида – субстратом s1. Область IV, границей которой является ломаная линия MNAPQ, – область нулевой биомассы сообщества. Коэффициенты угла наклона прямых, являющихся границами областей, есть ![]() и
и ![]() . Величины
. Величины ![]() на
на 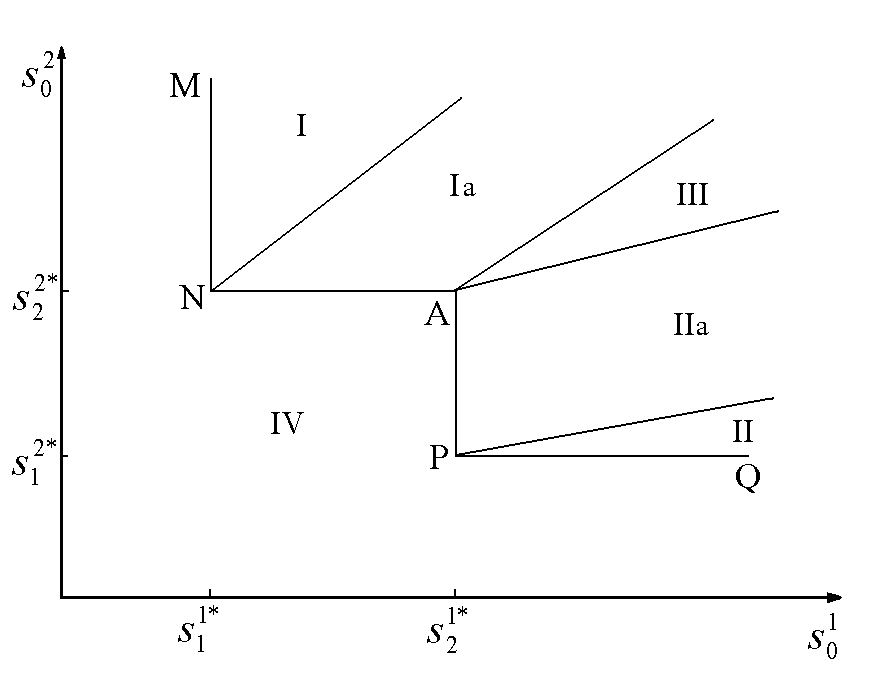 рис.7 есть концентрации j-го субстрата, при которых скорость роста i-го вида равна скорости его элиминации (m
i = D).
рис.7 есть концентрации j-го субстрата, при которых скорость роста i-го вида равна скорости его элиминации (m
i = D).
Рис.7. Разбиение плоскости начальных концентраций питательных веществ ![]() на области с разными режимами лимитирования и исходами конкуренции. В области I реализуется преимущество первого вида при лимитировании обоих конкурентов субстратом s1; в области Ia побеждает первый вид при лимитировании первого вида субстратом s2, а второго – субстратом s1; в области II конкуренцию выигрывает второй вид при лимитировании обоих конкурентов субстратом s2; в области IIa выигрывает второй вид при лимитировании второго вида субстратом s1, а первого вида – субстратом s2. В области III реализуется устойчивое сосуществование конкурентов при лимитировании первого вида субстратом s2, а второго вида – субстратом s1. Область IV, границей которой является ломаная линия MNAPQ, – область нулевой биомассы сообщества. Величины
на области с разными режимами лимитирования и исходами конкуренции. В области I реализуется преимущество первого вида при лимитировании обоих конкурентов субстратом s1; в области Ia побеждает первый вид при лимитировании первого вида субстратом s2, а второго – субстратом s1; в области II конкуренцию выигрывает второй вид при лимитировании обоих конкурентов субстратом s2; в области IIa выигрывает второй вид при лимитировании второго вида субстратом s1, а первого вида – субстратом s2. В области III реализуется устойчивое сосуществование конкурентов при лимитировании первого вида субстратом s2, а второго вида – субстратом s1. Область IV, границей которой является ломаная линия MNAPQ, – область нулевой биомассы сообщества. Величины ![]() – концентрации j-го субстрата, при которых скорость роста i-го вида равна скорости его элиминации.
– концентрации j-го субстрата, при которых скорость роста i-го вида равна скорости его элиминации.
Необходимо отметить, что при описании сообщества из w видов, потребляющих m ресурсов, авторы исходили из следующей формулировки принципа конкурентного исключения: “для равновесного сосуществования совокупности конкурирующих видов, не взаимодействующих друг с другом помимо потребления общих субстратов и не поедаемых хищниками, необходимо, чтобы число различных ресурсов было не меньше числа конкурентов” (Абросов, 1999). В соответствии с этим принципом в стационарной экосистеме не может быть более m видов. Пусть матрица приспособленностей имеет вид
 .
.
Тогда для сосуществования, например, первых m видов, которым соответствует квадратная матрица ![]() , должно выполняться условие
, должно выполняться условие  . Или – любая строка матрицы сообщества вытесненных видов должна иметь хотя бы один элемент, меньший минимального элемента соответствующего столбца матрицы приспособленности сообщества видов, выигравших конкуренцию (Абросов, 1999). Как отмечается в той же работе, практически все теоретические результаты получили качественное и количественное подтверждение для сообщества микроводорослей.
. Или – любая строка матрицы сообщества вытесненных видов должна иметь хотя бы один элемент, меньший минимального элемента соответствующего столбца матрицы приспособленности сообщества видов, выигравших конкуренцию (Абросов, 1999). Как отмечается в той же работе, практически все теоретические результаты получили качественное и количественное подтверждение для сообщества микроводорослей.
3.3.2. Эффекты влияния световой энергии. Кроме рассмотренных ситуаций лимитирования роста организмов питательными веществами, микроводоросли часто развиваются в условиях, когда их рост лимитируется световой энергией. Подробно были рассмотрены случаи лимитирования роста двух видов только световой энергией и лимитирования световой энергией и некоторым веществом (Абросов, Ковров, 1977). Для ситуации, когда все питательные вещества для двух видов, растущих в условиях хемостата, находятся в избытке, уравнения, описывающие динамику роста, имеют вид
 xi – концентрация клеток i-го вида; μi – удельная скорость роста i-го вида; D – скорость протока в хемостате;
xi – концентрация клеток i-го вида; μi – удельная скорость роста i-го вида; D – скорость протока в хемостате; ![]() – максимально возможная удельная скорость роста организмов i-го вида;
– максимально возможная удельная скорость роста организмов i-го вида; ![]() – процентное содержание хлорофилла в биомассе i-го вида; li – коэффициент поглощения света i-м видом;
– процентное содержание хлорофилла в биомассе i-го вида; li – коэффициент поглощения света i-м видом; ![]() – КПД фотосинтеза i-го вида клеток; Yi – величина, обратная калорийности биомассы i-го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора.
– КПД фотосинтеза i-го вида клеток; Yi – величина, обратная калорийности биомассы i-го вида клеток; E – мощность ФАР (фотосинтетически активной радиации), падающей на поверхность культуры; d – толщина слоя культуры в кювете культиватора.
Анализ системы показывает, что в культиваторе произойдет закрепление одного вида с максимальным коэффициентом приспособленности к световой энергии (![]() – коэффициент приспособленности i-го вида к световой энергии).
– коэффициент приспособленности i-го вида к световой энергии).
При возможном лимитировании роста двух видов световой энергией E и некоторым веществом s, модель имеет вид
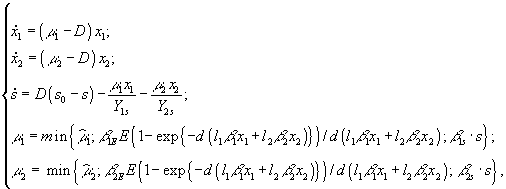
где s – концентрация ресурса в культуральной среде; s0 – концентрация вещества в поступающей питательной среде; ![]() – коэффициент приспособленности i-го вида к веществу s; Yis – экономический коэффициент использования вещества s i-м видом; остальные обозначения те же, что и в предыдущей формуле.
– коэффициент приспособленности i-го вида к веществу s; Yis – экономический коэффициент использования вещества s i-м видом; остальные обозначения те же, что и в предыдущей формуле.
Исследование данной системы позволяет построить разбиение пространства ![]() на области, в которых имеют место различные исходы конкуренции. Это разбиение зависит от вида функции
на области, в которых имеют место различные исходы конкуренции. Это разбиение зависит от вида функции ![]() . Результаты исследования этой модели аналогичны вышеприведенным результатам модели конкуренции двух видов за два ресурса, что позволяет описывать сообщество общей матрицей приспособленности, считая световую энергию одним из ресурсов.
. Результаты исследования этой модели аналогичны вышеприведенным результатам модели конкуренции двух видов за два ресурса, что позволяет описывать сообщество общей матрицей приспособленности, считая световую энергию одним из ресурсов.
Влияние интенсивности света на сосуществование видов микроводорослей исследовалось и другими авторами. В частности, А.Я.Болсуновским (1999) была предложена следующая математическая модель динамики компонентов сообщества из двух видов (спирулина и хлорелла), рост которых лимитировался интенсивностью света всей области ФАР (фотосинтетически активной радиации)
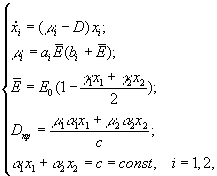
где xi – концентрация клеточного хлорофилла; μi – удельные скорости роста; ai, bi – константы уравнения Михаэлиса-Ментен; ![]() – средняя освещенность для всего сообщества; E0 – поверхностная освещенность.
– средняя освещенность для всего сообщества; E0 – поверхностная освещенность.
 В работе было построено разбиение пространства
В работе было построено разбиение пространства ![]() на области с различными исходами конкуренции (рис. 8).
на области с различными исходами конкуренции (рис. 8).
Рис.8. Области различных исходов конкуренции двух видов, рост которых лимитируется интенсивностью света Е0. В области I закрепляется второй вид (спирулина); в области III – первый (хлорелла); в области II – оба вида существуют совместно. Область IV – область, в которой данные монокультуры не существуют
В работе так же были исследованы модели: 1) два вида – лимитирование интенсивностью света отдельных спектральных областей; 2) три вида – лимитирование роста интенсивностью света отдельных спектральных областей ФАР; 3) два вида – ингибирование роста интенсивностью света всей области ФАР; 4) два вида – ингибирование роста одного вида и лимитирование роста другого вида интенсивностью света всей области ФАР; 5) два вида – лимитирование роста интенсивностью света всей области ФАР и наличие взаимодействия между видами через рассеянный световой поток. Для всех рассмотренных моделей теоретические расчеты получили подтверждение в экспериментах.
3.3.3. Перестройки структуры фитопланктонного сообщества. Изучение структуры фитопланктонного сообщества, состоящего из двух групп микроводорослей, лимитируемых одним биогенным элементом, предлагается в работе Ю.А.Домбровского с соавторами (1990). В качестве задачи ставится выявление условий, при которых могут происходить определенные структурные перестройки сообщества в результате увеличения лимитирующего ресурса. Отличительной чертой модели является учет регуляторного механизма меж- и внутривидовой конкуренции. Кинетика первичного круговорота описывается системой
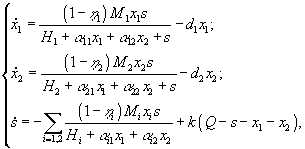 (3.6)
(3.6)
где x1, x2, s – концентрации двух видов фитопланктона и минерального вещества, выраженные в единицах лимитирующего биогенного вещества; ![]() – коэффициенты метаболизма;
– коэффициенты метаболизма; ![]() – относительные скорости отмирания водорослей; k – скорость минерализации органического вещества; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения;
– относительные скорости отмирания водорослей; k – скорость минерализации органического вещества; Mi – максимальная скорость фотосинтеза; Hi – параметр насыщения; ![]() – эмпирический коэффициент ингибирования i-го вида j-м; Q – запас биогенного вещества, участвующего в круговороте.
– эмпирический коэффициент ингибирования i-го вида j-м; Q – запас биогенного вещества, участвующего в круговороте.
Предполагается, что все коэффициенты положительны и выполняется необходимое условие живучести ![]()
Изучение вопроса об эволюции фитопланктонного сообщества при изменении ресурса биогенного элемента, участвующего в круговороте, дает три варианта зависимости стационарных концентраций двух сосуществующих видов фитопланктона от запаса ресурса (условие устойчивого сосуществования двух видов считается выполненным):
1) первый вид с самого начала захватывает лидерство и удерживает его в системе независимо от количества питательного вещества (рис.9а);
2) с ростом величины Q ситуация становится более благоприятной для второго вида, происходит смена доминирования (рис.9б);
3) первый вид, лучше приспособленный к минеральному субстрату, может полностью быть вытесненным более агрессивным, но нуждающимся в значительных количествах биогенных соединений вторым видом (рис.9в).
Математически и графически эти исходы выражаются следующим образом (для определенности считается, что ![]() , где
, где ![]() ).
).
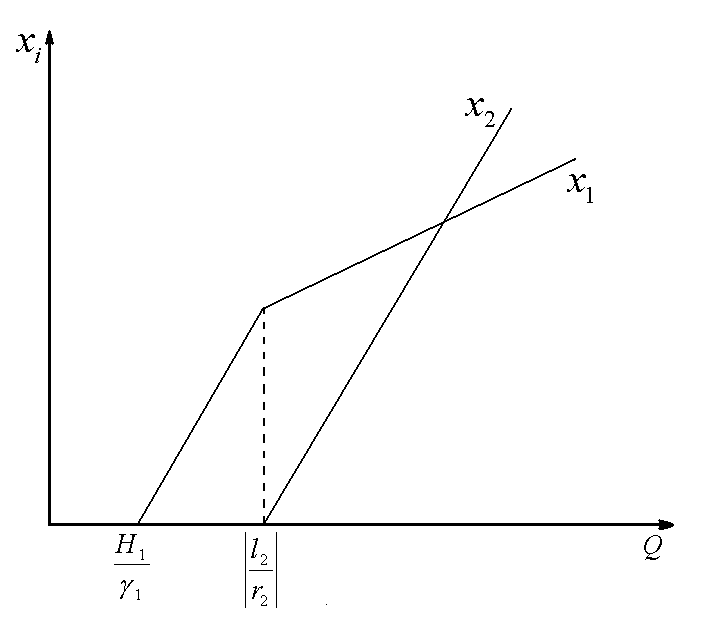
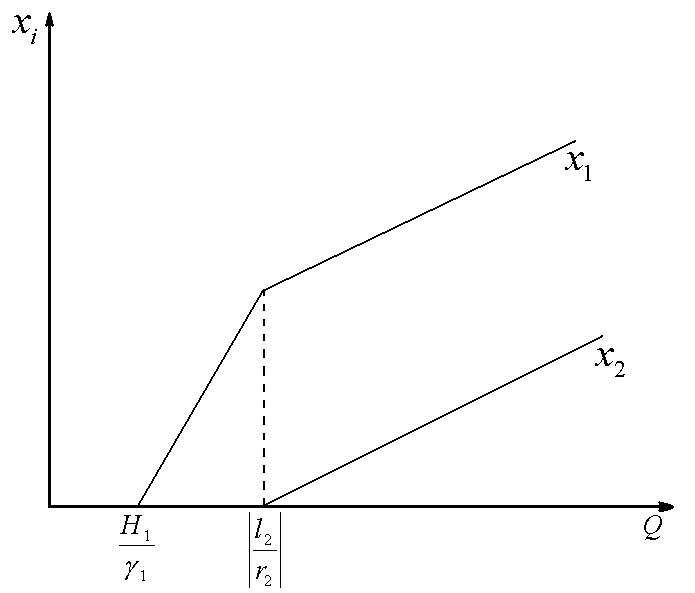 а) б)
а) б)

в)
Рис.9. Различные варианты эволюции фитопланктонного сообщества. x1, x2, – концентрации двух видов фитопланктона; Q – запас биогенного вещества, участвующего в круговороте. а) Первый вид с самого начала захватывает лидерство и удерживает его в системе независимо от количества питательного вещества; б) с ростом величины Q ситуация становится более благоприятной для второго вида, происходит смена доминирования; в) первый вид, лучше приспособленный к минеральному субстрату, может полностью быть вытесненным более агрессивным, но нуждающимся в значительных количествах биогенных соединений вторым видом
В формулах приняты обозначения
![]() ,
,
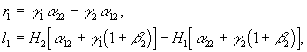
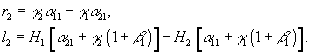
Случаи I и II принципиально не отличаются. При r1 > r2 имеет место ситуация, изображенная на рис. 9а); ситуация r1 < r2 представлена на рис. 9б); и случай III при r1 < 0 продемонстрирован на рис. 9в).
На основе качественного изучения системы (3.6) авторами были сделаны выводы:
1) “существование регуляторного механизма меж- и внутривидовой конкуренции, учтенного в модели (3.6), обеспечивает достаточное разнообразие структурных перестроек, сопутствующих процессам эвтрофирования”;
2) “возможный путь сукцессии фитопланктонного сообщества определяется не только экологическими особенностями отдельных видов, но и интенсивностью протекания совокупности деструкционных процессов в ходе круговорота органического вещества, характеризуемой параметром k”.
Видовой состав и сезонная сукцессия фитопланктона изучали в работе А.Доты (Dauta, 1983) на примере сообщества, состоящего из восьми видов водорослей. Клетки культуры, находящиеся в фазе экспоненциального роста, засевали в среду и наблюдали за их развитием при различных комбинациях температуры и освещенности. Полученные в экспериментах данные позволили разработать модель, в которой учитываются данные о пищевых ресурсах, температуре, освещенности, самозатенении, уровне ассимиляции и внутриклеточном накоплении питательных веществ.
Для описания изменений в популяции используют максимальные значения параметров роста ![]() , зависящие от коэффициентов торможения развития культур в случае неоптимальных условий – температуры (Т), освещенность (I) и наличия элементов питания (N, P, Si)
, зависящие от коэффициентов торможения развития культур в случае неоптимальных условий – температуры (Т), освещенность (I) и наличия элементов питания (N, P, Si)
![]() .
.
Здесь Red i – обозначает степень замедления роста под влиянием переменной i, а символ “*” означает комбинирование различных лимитирующих факторов, которое не обязательно осуществляется путем перемножения.
Наличие параметра освещения позволяет в неявной форме учитывать уровень метаболизма углерода (фотосинтез, дыхание).
Для выражения оптимального уровня роста, зависящего от освещенности, применяют уравнение Стила
![]() .
.
Ослабление освещенности (Ii) в результате самозатенения рассчитывают, исходя из закона Беера-Ламберта
![]() ,
,
где k – константа, определяемая геометрическими параметрами изучаемой культуры, Dopt – сумма относительных оптических плотностей, создаваемых клетками каждого вида водорослей.
Снижение скорости роста при обеднении среды описывают выражением
![]() ,
,
где ![]() и
и ![]() – внутриклеточное содержание питательного элемента j в момент времени t, минимальное и максимальное количество этого вещества, при котором рост прекращается, соответственно.
– внутриклеточное содержание питательного элемента j в момент времени t, минимальное и максимальное количество этого вещества, при котором рост прекращается, соответственно.
Скорость ассимиляции питательных веществ зависит от концентрации в среде sj элемента j и от внутриклеточных запасов

![]() ,
,
где ![]() – скорость ассимиляции питательных веществ;
– скорость ассимиляции питательных веществ; ![]() – максимальная скорость ассимиляции; Kj – константа полунасыщения j-м питательным веществом; x – число клеток; μ – скорость роста; остальные обозначения те же, что и в предыдущей формуле.
– максимальная скорость ассимиляции; Kj – константа полунасыщения j-м питательным веществом; x – число клеток; μ – скорость роста; остальные обозначения те же, что и в предыдущей формуле.
Количество хлорофилла в единичной клетке (qch) связано с внутриклеточными запасами питания элемента j соотношением
![]()
где ![]() ,
, ![]() – минимальное и максимальное количества хлорофилла, при которых рост останавливается, в случае недостаточности пищевого элемента j;
– минимальное и максимальное количества хлорофилла, при которых рост останавливается, в случае недостаточности пищевого элемента j; ![]() – внутриклеточное содержание питательного элемента j в момент времени t и минимальное количество этого вещества в клетке;
– внутриклеточное содержание питательного элемента j в момент времени t и минимальное количество этого вещества в клетке; ![]() – константы, рассчитываемые для каждого элемента.
– константы, рассчитываемые для каждого элемента.
Согласно результатам исследования, содержание хлорофилла (qch) в зависимости от запасов азота (qN) или фосфора (qP), описываются законом типа
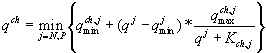 .
.
Проверяли пригодность модели для долгосрочного прогнозирования. В результате сравнения данных, полученных в экспериментах и предсказанных моделью, было обнаружено хорошее совпадение при описании следующих явлений: быстрое поглощение питательных веществ; поглощение света и запасание питательных веществ; дифференциальный рост относительного уровня ассимиляции; торможение роста в результате самозатемнения; снижение содержания хлорофилла в случаях крайнего ограничения в питании; сукцессия видов без учета ауто- и итероантогонизма.
Применение математических моделей с использованием дифференциальных уравнений возможно для описания такого явления как “цветение” воды. На основе уравнений Вольтерра С.В.Крестиным и Г.С.Розенбергом (1996) было предложено объяснение механизма этого явления. Рассматривали три группы популяций (“обычные”, синезеленые водоросли и зоопланктон), взаимодействие которых описывается системой
![]()
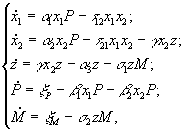
где x1, x2 – концентрации “обычных” и синезеленых водорослей; z – концентрация зоопланктона; ![]() и
и ![]() – коэффициенты экспоненциального роста водорослей, прямо пропорциональные концентрации фосфора P;
– коэффициенты экспоненциального роста водорослей, прямо пропорциональные концентрации фосфора P; ![]() – коэффициент экспоненциального вымирания зоопланктона, причем количество погибшего зоопланктона, кроме естественной смертности, прямо пропорционально концентрации зоопланктона и концентрации тяжелых металлов М;
– коэффициент экспоненциального вымирания зоопланктона, причем количество погибшего зоопланктона, кроме естественной смертности, прямо пропорционально концентрации зоопланктона и концентрации тяжелых металлов М;![]() и
и ![]() – коэффициенты межвидовой конкуренции;
– коэффициенты межвидовой конкуренции; ![]() – коэффициент переработки биомассы синезеленых водорослей в биомассу зоопланктона;
– коэффициент переработки биомассы синезеленых водорослей в биомассу зоопланктона; ![]() и
и ![]() – постоянное количество поступающего в воду фосфора и тяжелых металлов в единицу времени;
– постоянное количество поступающего в воду фосфора и тяжелых металлов в единицу времени; ![]() ,
, ![]() ,
, ![]() и
и ![]() – коэффициенты пропорциональности скоростей изменения биогенов и тяжелых металлов. Предполагалось, что
– коэффициенты пропорциональности скоростей изменения биогенов и тяжелых металлов. Предполагалось, что 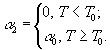 Температура определяется в каждой точке водоема по уравнению линейной регрессии. Авторами работы были исследованы стационарные решения данной системы, построены графики зависимостей x2(t), z(t), M(t) при двух наборах коэффициентов. Описан возможный механизм возникновения пиков на графиках с точки зрения взаимодействия синезеленых водорослей и зоопланктона. Предложенная авторами модель может служить основой для более детальной имитационной модели эвтрофирования равнинного водохранилища.
Температура определяется в каждой точке водоема по уравнению линейной регрессии. Авторами работы были исследованы стационарные решения данной системы, построены графики зависимостей x2(t), z(t), M(t) при двух наборах коэффициентов. Описан возможный механизм возникновения пиков на графиках с точки зрения взаимодействия синезеленых водорослей и зоопланктона. Предложенная авторами модель может служить основой для более детальной имитационной модели эвтрофирования равнинного водохранилища.
3.3.4. Конкурентное исключение или устойчивое сосуществование? Начиная с пионерских работ Г.Ф.Гаузе (Гаузе, 1935; Gause, 1935) и Г.Хатчинсона (Hatchinson, 1961) биологи и вслед за ними создатели математических моделей обсуждают проблему “Сколько видов может существовать на общих ресурсах?”.
Принцип конкурентного исключения ведет свое начало с работы В.Вольтерра (Volterra, 1928). В ней исследовалась система обыкновенных дифференциальных уравнений, описывающих конкуренцию двух видов за один ресурс
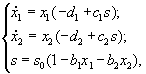 (3.7)
(3.7)
где xi – плотности вида i; s – колическтво ресурса; di – показатели смертности видов в отсутствии ресурса; s0 – начальное количество ресурса; положительные константы ci и bi , i = 1,2 характеризуют эффективность использования ресурса. Кроме того, предполагается выполненным условие ![]() , которое показывает, что при низких плотностях видов они растут за счет использования ресурса. После подстановки выражения для s в первые два уравнения системы (3.7) последняя принимает вид
, которое показывает, что при низких плотностях видов они растут за счет использования ресурса. После подстановки выражения для s в первые два уравнения системы (3.7) последняя принимает вид
 (3.8)
(3.8)
где ![]() .
.
В.Вольтерра показал, что независимо от начальных величин ![]() и
и ![]() при
при ![]() выигрывает вид с наибольшим значением
выигрывает вид с наибольшим значением ![]() , а численность другого вида стремится к нулевой величине. Если же
, а численность другого вида стремится к нулевой величине. Если же ![]() , то существует целая прямая положений равновесия
, то существует целая прямая положений равновесия ![]() , но этот случай также трактуется как очень маловероятный в реальности. Кроме того, в последнем случае ни одна из точек равновесия не будет ассимптотически устойчивой, что обычно требуется для биологических моделей.
, но этот случай также трактуется как очень маловероятный в реальности. Кроме того, в последнем случае ни одна из точек равновесия не будет ассимптотически устойчивой, что обычно требуется для биологических моделей.
Модель (3.7) – (3.8) легко обобщается на случай, когда w видов конкурируют за единственный ресурс. И в этой ситуации выигрывает вид с наибольшим значением ![]() .
.
Т.о. В.Вольтерра бал сделан вывод о том, что при конкуренции n видов за единственный ресурс, независимо от начальной численности видов, выживает лишь один вид.
Лабораторная проверка модели Вольтерра была проведена в сериях опытов Г.Ф.Гаузе (Гаузе, 1935; Gause, 1935) на двух видах одноклеточных, которые конкурировали за один ресурс. Подробно проанализировав эти опыты (Уильямсон, 1975; Барабашева и др., 1996 ), можно сделать вывод, что далеко не во всех опытах Г.Ф.Гаузе получено подтверждение выживания одного вида на одном ресурсе, а количественное согласие его результатов с решением системы уравнений (3.8) вообще практически нигде не было получено. (Стоит отметить, что современное осмысление работ Г.Ф.Гаузе приводит к выводу о принципиальной невозможности использовать уравнения вида ![]() для описания динамики численностей взаимодействующих видов (Тутубалин и др., 1997; 1999).)
для описания динамики численностей взаимодействующих видов (Тутубалин и др., 1997; 1999).)
Однако, результаты опытов Г.Ф.Гаузе были проинтерпретированы таким образом, что положение о том, что число неограниченно живущих вместе видов не может превышать число ресурсов, ограничивающих их существование, стало практически аксиомой и получило название "принципа конкурентного исключения", или "принципа Гаузе".
Существует очень большое число исследований конкуренции в естественных условиях, результаты которых сами авторы считают подтверждением принципа конкурентного исключения (см., например, ссылки в Levin, 1970). Однако почти все эти подтверждения носят косвенный характер. Например, Р.Мак Артур (Mc Artur, 1958) изучал пять видов певчих птиц, которые принадлежат к одному и тому же роду и, следовательно, очень близки в своих потребностях. В месте их обитания, они ограничены, в общем-то, только питанием и в некоторой степени пространством. Р.Мак Артур показал, что каждый вид занимает свой "уровень" леса, и расценил этот факт как подтверждение принципа Гаузе.
Опытное проявление принципа конкурентного исключения, как правило, наблюдается в экспериментах по проточному культивированию сообществ одноклеточных организмов, а математические модели, описывающие подобные ситуации, предсказывают строгое выполнение указанного принципа. Многочисленные опыты по непрерывному культивированию w видов организмов на одном ограничивающем рост ресурсе демонстрируют выживание лишь одного из видов, а именно, того, который способен существовать на самом низком уровне ресурса (см., например, Tilman,1982 и соответствующие ссылки). Адекватная модель для такой ситуации основана на модели Моно и имеет вид (Hsu et. al., 1977):
 (3.9)
(3.9)
с начальными условиями ![]() Здесь xi(t) – концентрация i-го вида; s(t) – концентрация субстрата, лимитирующего рост;
Здесь xi(t) – концентрация i-го вида; s(t) – концентрация субстрата, лимитирующего рост; ![]() обозначает максимально возможную удельную скорость роста организмов i-го вида; Yi-1 – "экономический коэффициент" i-го вида; s0 – концентрация субстрата во внешнем резервуаре; D – скорость протока; Ki. – константа половинного роста i-го вида.
обозначает максимально возможную удельную скорость роста организмов i-го вида; Yi-1 – "экономический коэффициент" i-го вида; s0 – концентрация субстрата во внешнем резервуаре; D – скорость протока; Ki. – константа половинного роста i-го вида.
Результаты исследования (3.9) следующие:
 ,
, Таким образом, в этой модели имеет место классическая ситуация конкурентного исключения.
В работах А.Г.Дегерменджи (1981; 1986) рассмотрена модель роста w видов в открытой системе типа хемостата, удельная скорость каждого вида зависит от m факторов среды, которые, в свою очередь, находятся под контролем плотностей указанных видов. Такая модель описывается уравнениями
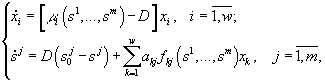 (3.10)
(3.10)
где xi – плотность i-го вида; ![]() – удельная скорость роста; D – скорость протока;
– удельная скорость роста; D – скорость протока; ![]() – концентрации j-го ресурса на входе и в среде; член
– концентрации j-го ресурса на входе и в среде; член ![]() определяет скорость выработки или поглощения j-го вещества k-м видом. Доказано, что возможное число стационарно сосуществующих видов не превышает числа контролирующих рост независимых факторов, определяемых плотностями этих видов (
определяет скорость выработки или поглощения j-го вещества k-м видом. Доказано, что возможное число стационарно сосуществующих видов не превышает числа контролирующих рост независимых факторов, определяемых плотностями этих видов (![]() ). Данное утверждение верно и для смешанной культуры, в которой существует предельный цикл (устойчивый или нет) с периодом Т, с учетом предположения о линейности функций
). Данное утверждение верно и для смешанной культуры, в которой существует предельный цикл (устойчивый или нет) с периодом Т, с учетом предположения о линейности функций ![]() и
и ![]() . При рассмотрении вместо второго уравнения системы (3.10) более общего
. При рассмотрении вместо второго уравнения системы (3.10) более общего
![]()
“запрет” на избыточное число сосуществующих видов сохраняется (в этом уравнении B1,...,Bk – концентрации факторов, влияющих на динамику j-го фактора, но не влияющих на динамику видов; функции Ψj отражают влияние на динамику концентраций j-го фактора химических реакций между факторами, деградации этого фактора, его синтеза через некоторые другие промежуточные вещества B1,...,Bk и других подобных явлений).
Другая частная формулировка принципа конкурентного исключения была предложена А.Г.Боголюбовым (1995). Автором рассмотрена модель
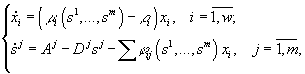
где xi – плотность популяции вида i; sj – плотность ресурса j; ![]() и
и ![]() – удельная скорость роста и элиминации популяции i; Aj и Dj – скорость поступления и удельная скорость удаления или распада ресурса j;
– удельная скорость роста и элиминации популяции i; Aj и Dj – скорость поступления и удельная скорость удаления или распада ресурса j; ![]() – удельная скорость потребления ресурса j видом i. Если 1) виды конкурируют и только конкурируют друг с другом; 2) ни один вид не производит ингибиторов или стимуляторов своего роста и возобновления; 3) удельные скорости роста видов не зависят от плотностей популяций; 4) удельные скорости элиминации популяций не зависят от их плотностей; 5) скорости поступления ресурсов – постоянные положительные величины; 6) все ресурсы не взаимодействуют друг с другом; 7) физические условия окружающей среды постоянны во времени и не выходят за пределы толерантных диапазонов конкурирующих видов; 8) скорости поступления ресурсов таковы, что рост и возобновление всех конкурирующих видов лимитируется единственным и общим для всех трофическим ресурсом, тогда их сосуществование невозможно, и из всего первоначального сообщества выживает не более одного вида. Ослабление некоторых из этих условий приводит к отрицанию принципа конкурентного исключения.
– удельная скорость потребления ресурса j видом i. Если 1) виды конкурируют и только конкурируют друг с другом; 2) ни один вид не производит ингибиторов или стимуляторов своего роста и возобновления; 3) удельные скорости роста видов не зависят от плотностей популяций; 4) удельные скорости элиминации популяций не зависят от их плотностей; 5) скорости поступления ресурсов – постоянные положительные величины; 6) все ресурсы не взаимодействуют друг с другом; 7) физические условия окружающей среды постоянны во времени и не выходят за пределы толерантных диапазонов конкурирующих видов; 8) скорости поступления ресурсов таковы, что рост и возобновление всех конкурирующих видов лимитируется единственным и общим для всех трофическим ресурсом, тогда их сосуществование невозможно, и из всего первоначального сообщества выживает не более одного вида. Ослабление некоторых из этих условий приводит к отрицанию принципа конкурентного исключения.
Согласно принципа конкурентного исключения количество сосуществующих видов не может превышать количества факторов, лимитирующих их рост. Однако в природе наблюдаются ситуации, противоречащие этому принципу. В частности, явление сосуществования большого числа видов фитопланктона было названо “парадоксом фитопланктона”. Этот термин впервые употребил Г.Хатчинсон (Hatchinson, 1961), выдвинувший предположение о том, что небольшие различия в температурных нишах и прочих факторах среды приводят к чередованию периодов доминирования различных видов фитопланктона в переменных внешних условиях (Домбровский и др., 1990). Было высказано много гипотез, объясняющих наблюдаемое противоречие, которые можно разделить на две группы – равновесные и неравновесные модели (Gonzalez, 1997). В неравновесных моделях фактором, вызывающим нарушение принципа конкурентного исключения, является временная и пространственная неоднородность. Временные неоднородности создаются благодаря внутренним взаимодействиям между компонентами экосистемы при постоянных внешних характеристиках среды, например, колебательное функционирование, присущее взаимодействию “хищник-жертва” или происходящее благодаря флуктуациям внешних по отношению к экосистеме параметров, например, сезонные колебания обеспеченности ресурсами, температуры и др. (Абросов, 1999). Примером может служить модель Ф.Н.Стюарта и Б.Р.Левина, описанная в работе А.Г.Дегерменджи (1981). В модели рассмотрена конкуренция двух видов, лимитированных одним ресурсом, растущих в режиме периодического уменьшения численности в d раз и восстановления ресурса до концентрации R
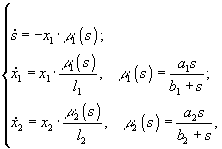
где s – доступное количество ресурса; x1, x2 – численности видов; l1, l2 – коэффициенты потребления; μ1(s), μ2(s) – удельные скорости роста; ai, bi – кинетические параметры. Были получены условия сосуществования двух видов и показано, что бывают такие наборы кинетических параметров и других постоянных (d, R, l1, l2), при которых эти условия выполняются.
В общем случае была предложена модель сосуществования любого числа конкурирующих видов на одном ресурсе в нестационарной пространственной среде. Авторы этой модели Р.А.Армстронг и Р.МакГечи рассматривали w видов, динамика которых описывается системой (Дегерменджи, 1981):
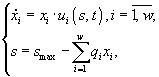
где xi – биомасса вида i; функция ui отражает особенности роста вида i; smax – максимальное количество ресурса, которое может быть использовано видами; qi – количество вещества, потребляемое видом i.
Предполагается, что каждый вид имеет сезоны роста, отличные от других. Вне этого сезона каждый вид достаточно быстро “гибнет”, так что ростовой сезон “следующего” вида наступает при достатке корма s, т.е. виды слабо взаимодействуют друг с другом, практически не конкурируя. Результатом исследования данной модели является возможность динамического сосуществования w видов на одном ресурсе.
Важно отметить, что для сосуществования видов в переменных условиях нужно, чтобы существовал временной интервал для каждого вида, на котором этот вид имеет конкурентное преимущество перед остальными. Это замечание позволяет высказать утверждение о том, что устойчиво могут сосуществовать столько видов, для скольких возможна реализация преимущества перед другими видами, причем формулировка принципа конкурентного исключения будет являться частным случаем данного утверждения (Абросов, 1999).
Свой вклад в сосуществование видов вносит и пространственная неоднородность. В частности, при рассмотрении двух внутренне гомогенных хемостатов, связанных обменными потоками, в которых две конкурирующие популяции лимитированы общим потоком субстрата, было показано, что возможно сосуществование, но при относительно малом уровне обменных потоков (Абросов, 1999).
С целью объяснить одновременно эффекты видового разнообразия и “пятнистости” фитопланктона была высказана гипотеза, проверенная на математической модели, которая основывалась на предположении, что хорошо перемешиваемая вода существует в естественных условиях в течение коротких периодов в небольших “пятнах”, каждое их которых характеризуется одновидовым цветением планктона. Периодически, благодаря турбулентности, “пятна” разбиваются. Так создается планктонный ансамбль (Домбровский и др., 1990).
В качестве примера равновесной модели можно рассмотреть модель Р.Петерсена (Gonzalez, 1997). В рамках его гипотезы предполагаются выполненными условия:
1) некоторые питательные вещества относительно ограничены;
2) рост каждого вида лимитируется единственным ресурсом или уникальной комбинацией нескольких;
3) различные виды обладают различными способностями поглощать питательные вещества.
Ключевой элемент модели Р.Петерсена – существование равновесной концентрации лимитирующего питательного вещества ![]() , а именно, количества вещества j, необходимого для вида i, чтобы его скорость роста равнялась скорости отмирания; здесь
, а именно, количества вещества j, необходимого для вида i, чтобы его скорость роста равнялась скорости отмирания; здесь ![]() – максимальная скорость роста i-го вида; Kij – константа полунасыщения для вида i, лимитируемого веществом j; di – скорость смерти i-го вида.
– максимальная скорость роста i-го вида; Kij – константа полунасыщения для вида i, лимитируемого веществом j; di – скорость смерти i-го вида.
В условиях равновесия виды с наименьшим значением равновесной концентрации питательного вещества R* вытеснят другие виды.
Для двух конкурирующих видов предлагается графическое представление возможных исходов конкуренции (рис. 10). Границы областей на рис. 10 определяются потреблением видами питательных веществ s1 и s2.
Поскольку в обоих видах моделей существуют условия, при которых возможно сосуществование конкурирующих видов, было высказано предположение, что модель, использующая элементы и того, и другого подходов, окажется более подходящей для объяснения “парадокса фитопланктона” (Gonzalez, 1997).
 Рис.10. Возможные исходы конкуренции в модели Р.Петерсена (Gonzalez, 1997). В области I выигрывает второй вид, в области II – первый, в области III реализуется сосуществование видов. Kij – константа полунасыщения для вида i, лимитируемого веществом j; s1, s2 – концентрации питательных веществ в среде
Рис.10. Возможные исходы конкуренции в модели Р.Петерсена (Gonzalez, 1997). В области I выигрывает второй вид, в области II – первый, в области III реализуется сосуществование видов. Kij – константа полунасыщения для вида i, лимитируемого веществом j; s1, s2 – концентрации питательных веществ в среде
В книге Ю.А.Домбровского и его коллег (1990) приведена модель, с помощью которой можно объяснить явление “парадокса фитопланктона” на основе экспериментальных наблюдений метаболитного механизма регуляции численностей популяций планктонных водорослей. Динамика двух фитопланктонных популяций, с учетом механизма метаболитного регулирования, описывается системой уравнений

где ![]() – удельные скорости роста первого и второго видов; d1, d2 – коэффициенты смертности, γ – коэффициент экскреции метаболита M фитопланктоном, d3 – скорость разложения метаболита.
– удельные скорости роста первого и второго видов; d1, d2 – коэффициенты смертности, γ – коэффициент экскреции метаболита M фитопланктоном, d3 – скорость разложения метаболита.
Полагается, что влияние концентрации метаболита на рост фитопланктона описывается линейной зависимостью с параметрами ![]() и
и ![]() для первого и второго видов.
для первого и второго видов.
Знак параметра ![]() отражает характер воздействия метаболита (
отражает характер воздействия метаболита (![]() – стимулирующее воздействие,
– стимулирующее воздействие, ![]() – ингибирующее).
– ингибирующее).
Система имеет невырожденное состояние равновесия
Стационарные численности положительны, если:
1) ![]() и
и ![]() , т.е. первый вид имеет более высокие продукционные характеристики, но воздействие метаболита более благоприятно для второго вида;
, т.е. первый вид имеет более высокие продукционные характеристики, но воздействие метаболита более благоприятно для второго вида;
2) ![]() и
и ![]() , т.е. коэффициент приспособленности выше у второго вида, но метаболит оказывает на него более сильное ингибирующее (или более слабое стимулирующее) влияние, чем на первый.
, т.е. коэффициент приспособленности выше у второго вида, но метаболит оказывает на него более сильное ингибирующее (или более слабое стимулирующее) влияние, чем на первый.
Исследование устойчивости состояния равновесия показывает, что ситуация (1) дает стабильное сосуществование видов. Это состояние реализуется, если величина параметра ![]() не слишком велика.
не слишком велика.
Модель легко обобщается на случай произвольного числа видов фитопланктона, конкурирующих за единственный ресурс, причем можно доказать, что для устойчивого сосуществования w видов достаточно набора из (![]() ) метаболитов.
) метаболитов.
С помощью численных расчетов было проведено сопоставление приведенной выше модели с аналогичной, но не учитывающей метаболитного воздействия. Результат позволяет говорить, что с помощью гипотезы о наличии у фитопланктона механизма эктокринного регулирования можно объяснить парадокс видового разнообразия фитопланктона. Так, устойчивое сосуществование любого числа видов, конкурирующих за единственный ресурс, наблюдается в случае, когда происходит экскреция стимулирующих метаболитов высокопродуктивными видами, а также при выделении метаболита, стимулирующего рост других видов популяцией фитопланктона с более высокой приведенной скоростью роста.
С принципом конкурентного исключения Гаузе тесно связана концепция экологической ниши. Существует несколько совершенно разных определений данного понятия. В книге Ю.М.Свирежева и Д.О.Логофета (1978) был описан компроимссный вариант, допускающий возможность формализации. Пусть ресурс, потребляемый биологическими видами, характеризуется параметром s, а количество доступного для потребления ресурса с данными характеристиками определяется некоторой функцией K(s). Тогда множество значений s с заданной на нем функцией K(s) называется спектром ресурса. Экологическая ниша вида определяется точкой s0 на спектре ресурса и заданной функцией потребления ресурса данным видом (вероятностным распределением плотности f(s) со средним значением s0 и конечной дисперсией σ2). Конкуренция, связанная с перекрыванием экологических ниш конкурирующих видов, является результатом пересечения областей определения соответствующих функций потребления fi(s) в спектре ресурса (индекс i нумерует виды). Авторами был получен критерий стабильности конкурентного сообщества: выполнение условия существования положительного равновесия, т.е. положительного решения линейной системы
![]() ,
,
где ![]() – общее количество ресурса, потребленного i-м видом;
– общее количество ресурса, потребленного i-м видом; ![]() – коэффициент конкуренции между i-м и j-м видом, пропорциональный полной вероятности столкновения потребительских интересов данной пары видов в одной и той же точке спектра и характеризующий степень перекрывания экологических ниш;
– коэффициент конкуренции между i-м и j-м видом, пропорциональный полной вероятности столкновения потребительских интересов данной пары видов в одной и той же точке спектра и характеризующий степень перекрывания экологических ниш; ![]() – численности видов; w – число видов сообщества.
– численности видов; w – число видов сообщества.
В начале 80-х годов была высказана иная точка зрения на проблему конкурентного исключения. Так, авторы некоторых работ (например, Birch, 1979; Boer, 1980; Ghilarov, 1984), подробно проанализировав ряд случаев сосуществования, показали, что существуют примеры, где традиционные "простые" объяснения отсутствия конкурентного исключения не проходят, а объяснения, сделанные в рамках принципа Гаузе, настолько сложны и громоздки, что очень маловероятны, чтобы они имели какое-то отношение к реальности. В связи с этим Л.Бирч (Birch, 1979) пишет, что "в естественных популяциях, в противоположенность гипотетическим, исключение одного вида другим есть лишь экстремальный случай в широком диапазоне эффектов воздействия одного вида на другой, простирающемся от нулевого воздействия до полного исключения". Д.Боэр (Boer, 1980) выдвигает альтернативный "принцип сосуществования", как обобщение многочисленных эмпирических данных.
Таким образом, можно подвести следующий итог. Принцип конкурентного исключения, гласящий, что число неограниченно живущих видов не может превышать число ресурсов, ограничивающих их существование, есть лишь далеко идущее теоретическое обобщение результатов, полученных при исследовании модели очень специального вида. Поэтому этот принцип нужно рассматривать как некоторое правило, применимое к достаточно узкому кругу ситуаций, но не как действующую всюду аксиому.
3.3.5. “Графическая” теория межвидовой борьбы. Вопросами конкурентной борьбы за средства существования и изучением структуры сообщества занимался Д.Тилман (Tilman, 1982). В своей монографии он изложил “графическую” теорию межвидовой борьбы, в рамках которой аналитическое исследование систем дифференциальных и алгебраических уравнений заменено геометрической интерпретацией.
Ключевым понятием теории Д.Тилмана является средство существования, которым считается любое вещество или фактор, приводящий к ускорению роста по мере увеличения его количества в окружающей среде, и который потребляется организмом. В случае, когда какой-то вид потенциально лимитируется не одним, а несколькими ресурсами, фактор будет считаться средством существования, когда имеется какой-то диапазон наличия или доступности другого лимитирующего фактора, в котором первый удовлетворяет требованиям данного выше определения. Как следствие – возможность классифицирования средств существования на основе характера взаимодействия различных факторов, определяющих скорость роста какого-либо вида.
В основе классификации лежат общие уравнения, которые показывают, как средства существования влияют на рост популяции и как потребители влияют на количества средств существования

где xi – плотность популяции вида i; sj – запасы средства существования j; di – показатель смертности вида i; fi – функция, описывающая зависимость скорости размножения (в расчете на одну особь) от количества средства существования; gj– функция, характеризующая скорость поступления средства существования j; hij – функция, описывающая количество средства j, требующееся для создания каждой новой особи вида i; всего w видов конкурируют за m средств сущестовования.
В этих уравнениях помимо очевидных (непрерывное размножение, гомогенность популяций и мест обитания и др.) сделан ряд важных допущений: разные виды взаимодействуют между собой только через пользование средствами существования; между средствами существования нет никакого взаимодействия;
Д.Тилман выделяет восемь классов, охватывающих все основные формы реакций вида на два лимитирующих средства существования. Для демонстрации потребностей отдельных видов в этих средствах применяются “изоклины рост – средства существования”, отражающие все комбинации наличных средств существования, по которым у данного вида имеется заданная удельная скорость роста ![]() .
.
На рис.11 а) – з) представлены различные формы, принимаемые изоклинами роста в предположении, что данный вид обитает в местности, где отсутствует смертность, т.е. di = 0. Группа рисунков а) – г) представляет взаимозаменяемые ресурсы, т.е. каждый из них способен поддерживать размножение при отсутствии второго (на графиках это отражается пересечением изоклин с осями координат).
а) Полностью взаимозаменяемые ресурсы, одно средство можно заменить пропорциональным количеством другого;
в) антагонистические средства существования – когда данный вид потребляет одновременно оба средства существования, то для сохранения заданной скорости роста, ему требуется пропорционально больше ресурсов, чем при их потреблении по отдельности;
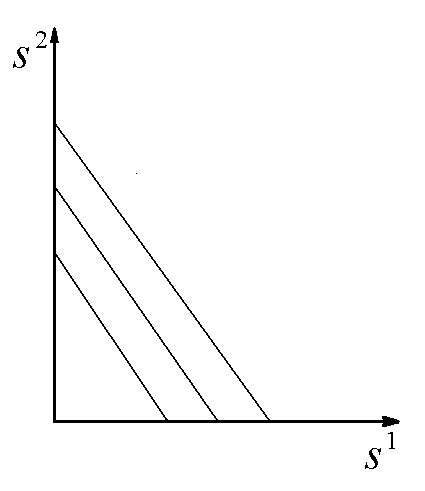
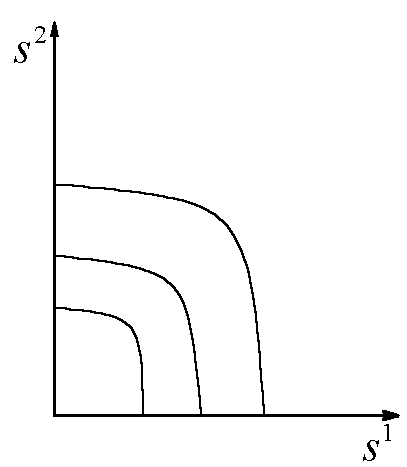
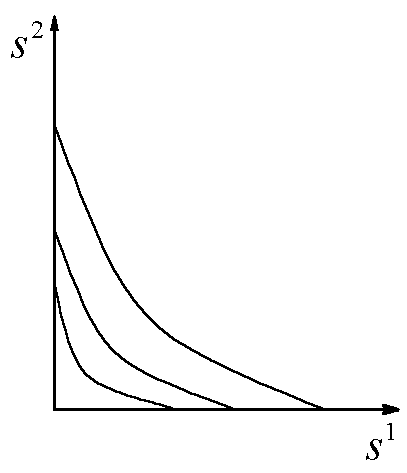
а) б) в)
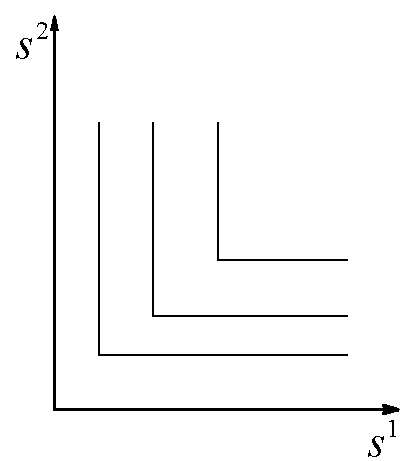
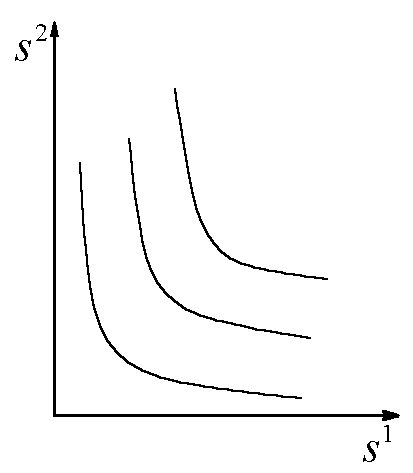

г) д) е)

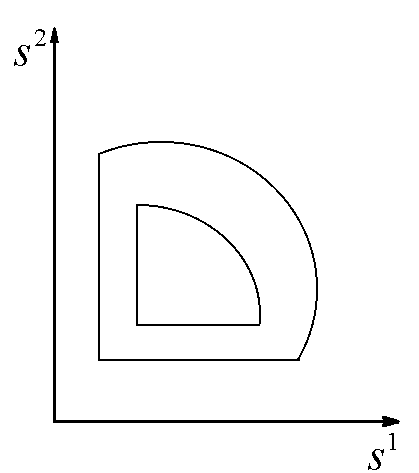
ж) з)
Рис.11. Изоклины роста организмов, потребляющих различные типы средств существования. sj – запасы средств существования в данной местности. а) Полностью взаимозаменяемые ресурсы, одно средство можно заменить пропорциональным количеством другого; б) комплементарные средства существования – при одновременном потреблении обоих ресурсов виду требуется меньшее количество средств, чем для прямых изоклин; в) антагонистические средства существования – когда данный вид потребляет одновременно оба средства существования, то для сохранения заданной скорости роста, ему требуется пропорционально больше ресурсов, чем при их потреблении по отдельности; г) полностью антагонистические средства (переключающиеся) – данный вид потребляет либо одно средство существования, либо второе, но не потребляет оба средства одновременно; д) полностью незаменимые средства существования – вид лимитирован одним или вторым средством, и полностью отсутствует взаимозаменяемость между этими средствами; е) взаимодействующие незаменимые средства – есть зоны количеств обоих средств, в которых одно средство может частично заменить другое; ж) полунезаменимые средства существования – одно совершенно необходимо для роста (изоклина пересекает ось, соответствующую этому ресурсу), а второе может отчасти заменить первое (изоклина проходит параллельно оси этого средства); з) изоклины образуют замкнутую кривую, что отражает ситуацию, когда при увеличении количества средств существования наступает момент уменьшения роста популяции
г) полностью антагонистические средства (переключающиеся) – данный вид потребляет либо одно средство существования, либо второе, но не потребляет оба средства одновременно.
На рисунках д) и е) представлены изоклины для средств существования, которые необходимы для роста и незаменимы, т.е. если какого-то ресурса мало, то второй не может его полностью заменить (графически этот факт отражается параллельностью изоклин осям).
д) полностью незаменимые средства существования – вид лимитирован одним или вторым средством, и полностью отсутствует взаимозаменяемость между этими средствами;
е) взаимодействующие незаменимые средства – есть зоны количеств обоих средств, в которых одно средство может частично заменить другое.
Последний класс – частично незаменимые средства существования (рис. ж) – одно совершенно необходимо для роста (изоклина пересекает ось, соответствующую этому ресурсу), а второе может отчасти заменить первое (изоклина проходит параллельно оси этого средства).
На последнем рисунке (рис. з) представлены изоклины, образующие замкнутую кривую, отражающую ситуацию, когда при увеличении количества средств существования наступает момент уменьшения роста популяции (торможение, угнетение роста).
Описанные изоклины роста являются одним из главных инструментов графического подхода.
Большая часть работы Д.Тилмана посвящена вопросам, связанным с разработкой приложения его метода к теории равновесия в конкуренции за два органических ресурса. Для прогнозирования равновесного положения результата конкуренции необходимы четыре элемента информации: скорость роста каждого вида, смертность каждого вида, темпы восстанавливаемости ресурсов и скорость потребления каждого ресурса каждым из видов. Состояние равновесия наступает, когда скорость роста равна смертности, а показатель восстанавливаемости ресурса равен показателю общего потребления для каждого ресурса. Половину информации, требующейся для определения положения равновесия, можно получить с помощью изоклины скорости роста, на которой величина показателя скорости роста для каждого вида уравнивается с величиной показателя смертности для этого вида (так называемая изоклина полного отсутствия роста). Изоклина полного отсутствия роста разделяет плоскость ресурсов ![]() на две части, в одной из которых плотность популяции вида будет уменьшаться, а в другой – увеличиваться. На рис. 12 а) – г) представлены изоклины полного отсутствия роста в случае: а) незаменимых ресурсов, б) дополнительных (комплементарных), в) заменимых, г) чередующихся.
на две части, в одной из которых плотность популяции вида будет уменьшаться, а в другой – увеличиваться. На рис. 12 а) – г) представлены изоклины полного отсутствия роста в случае: а) незаменимых ресурсов, б) дополнительных (комплементарных), в) заменимых, г) чередующихся.
Показатель потребления ресурсов может быть представлен вектором потребления ![]() , который берет свое начало из точки на изоклины полного отсутствия роста и является суммой двух компонентов – темпов общего потребления видом ресурсов s1 и s2
, который берет свое начало из точки на изоклины полного отсутствия роста и является суммой двух компонентов – темпов общего потребления видом ресурсов s1 и s2  . Здесь
. Здесь ![]() – показатель темпов потребления ресурса s1, в расчете на одного представителя i-го вида при равновесном состоянии;
– показатель темпов потребления ресурса s1, в расчете на одного представителя i-го вида при равновесном состоянии; ![]() – показатель темпов потребления ресурсов s2, взятый на одного представителя вида i при равновесном состоянии;
– показатель темпов потребления ресурсов s2, взятый на одного представителя вида i при равновесном состоянии; ![]() – плотность вида i при равновесном состоянии.
– плотность вида i при равновесном состоянии.
а) б)


в) г)
Рис.12. Области увеличения (темная область) и уменьшения (светлая область) плотности популяции, на которые изоклина полного отсутствия роста делит пространство средств существования. sj – запасы средств существования в данной местности. а) Незаменимый ресурс; б) дополнительный (комплементарный) ресурс; в) заменимый ресурс; г) чередующийся ресурс
Восстанавливаемость ресурсов представляется вектором ![]() , складывающимся из темпов восстанавливаемости ресурсов s1 и s2. Пусть Sj – количество ресурса j, которое может существовать в месте обитания при отсутствии потребления, тогда, предположив, что темпы восстанавливаемости ресурса являются пропорциональными величине (Sj – sj), т.е.
, складывающимся из темпов восстанавливаемости ресурсов s1 и s2. Пусть Sj – количество ресурса j, которое может существовать в месте обитания при отсутствии потребления, тогда, предположив, что темпы восстанавливаемости ресурса являются пропорциональными величине (Sj – sj), т.е. ![]() , можно определить вектор восстанавливаемости ресурсов
, можно определить вектор восстанавливаемости ресурсов  (постоянные темпов восстановления aj считаются равными a для всех ресурсов) (рис.13).
(постоянные темпов восстановления aj считаются равными a для всех ресурсов) (рис.13).

Рис.13. Изоклина полного отсутствия роста и векторы восстанавливаемости ресурсов ![]() для разных мест обитания А, В, С (с разной доступностью ресурсов), имеющих одну и ту же точку восстановления ресурсов (S1, S2)
для разных мест обитания А, В, С (с разной доступностью ресурсов), имеющих одну и ту же точку восстановления ресурсов (S1, S2)
Теперь известна вся необходимая информация для определения равновесного состояния. Из всех точек на изоклине полного отсутствия роста, существует только одна точка, в которой вектор потребления ресурса будет иметь направление, противоположное направлению вектора восстанавливаемости ресурса. Эта точка и будет точкой равновесного состояния (рис.14).
В случае конкуренции за два ресурса возможны четыре качественно различных случая равновесного состояния, которые проиллюстрированы с использованием изоклины полного отсутствия роста для незаменимых ресурсов (рис.15а) – г)):


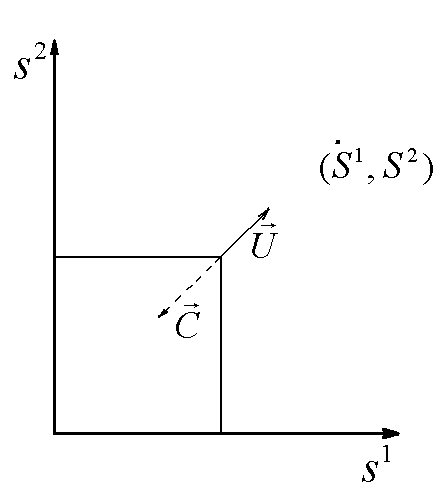 а) б)
а) б)
в)
Рис.14. Примеры расположения точки равновесного состояния по отношению к точке восстанавливаемости ресурсов (S1, S2) для некоторых классов ресурсов. ![]() – вектор восстанавливаемости ресурсов;
– вектор восстанавливаемости ресурсов; ![]() – вектор потребления. а) Незаменимый ресурс; б) дополнительный (комплементарный) ресурс; в) чередующийся ресурс
– вектор потребления. а) Незаменимый ресурс; б) дополнительный (комплементарный) ресурс; в) чередующийся ресурс
а) Виду А требуется меньшее количество обоих ресурсов для достижения величины показателя роста, равной величине показателя смертности, чем виду В. Точки восстанавливаемости ресурсов, находящиеся в области 1, характеризуют места с недостаточным количеством ресурсов для выживания любого вида, оба вида исчезнут. Места обитания с точкой восстанавливаемости из области 2 обладают ресурсами, достаточными для выживания вида А, и недостаточными для выживания вида В. В таком месте обитания вид А достигнет устойчивого равновесного состояния плотности популяции, а вид В исчезнет. В области 3 ресурсов достаточно для раздельного существования обоих видов, однако, при совместном существовании плотность популяции А будет увеличиваться до тех пор, пока точка восстанавливаемости ресурсов не сместится на изоклину полного отсутствия роста и вид В исчезнет.
б) Виды А и В меняются местами по сравнению со случаем (а).
в) Точка пересечения изоклин – точка равновесного состояния обоих видов, обладающая локальной устойчивостью (каждый из видов потребляет большее количество того ресурса, который в большей степени ограничивает его собственное развитие). Область 4 – сосуществование двух видов; области 2, 3 – доминирование вида В; область 1 – вымирание обоих видов.
г) точка пересечения – точка равновесного состояния, не обладающая локальной устойчивостью (каждый из видов потребляет в больших количествах тот тип ресурсов, который в меньшей степени ограничивает скорость его роста). Результаты конкуренции идентичны результатам случая в), за исключением того, что в области 4* верх одерживает либо вид А, либо вид В, в зависимости от начальных условий.



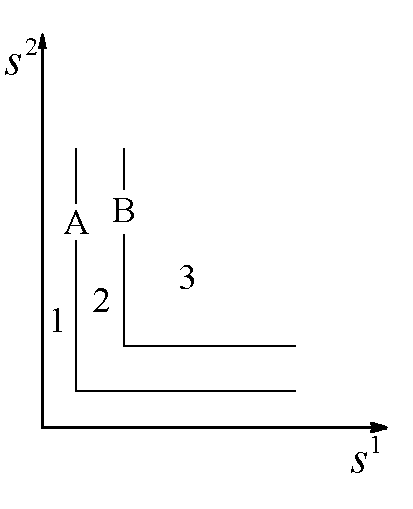 а) б)
а) б)
в) г)
Рис.15. Возможные исходы конкурентной борьбы видов А и В за ресурсы s1 и s2. ![]() – вектор потребления. а) В местах обитания с точкой восстанавливаемости, находящейся в области 1, оба вида исчезнут. В местах обитания с точкой восстанавливаемости из области 2 вид А достигнет устойчивого равновесного состояния плотности популяции, а вид В исчезнет. В области 3 ресурсов достаточно для раздельного существования обоих видов, однако, при совместном существовании плотность популяции А будет увеличиваться до тех пор, пока точка восстанавливаемости ресурсов не сместится на изоклину полного отсутствия роста и вид В исчезнет; б) виды А и В меняются местами по сравнению со случаем (а); в) точка пересечения изоклин – точка равновесного состояния обоих видов, обладающая локальной устойчивостью. Область 4 – сосуществование двух видов; области 2, 3 – доминирование вида В; область 1 – вымирание обоих видов; г) точка пересечения – точка равновесного состояния, не обладающая локальной устойчивостью. Результаты конкуренции идентичны результатам случая в), за исключением того, что в области 4* верх одерживает либо вид А, либо вид В в зависимости от начальных условий.
– вектор потребления. а) В местах обитания с точкой восстанавливаемости, находящейся в области 1, оба вида исчезнут. В местах обитания с точкой восстанавливаемости из области 2 вид А достигнет устойчивого равновесного состояния плотности популяции, а вид В исчезнет. В области 3 ресурсов достаточно для раздельного существования обоих видов, однако, при совместном существовании плотность популяции А будет увеличиваться до тех пор, пока точка восстанавливаемости ресурсов не сместится на изоклину полного отсутствия роста и вид В исчезнет; б) виды А и В меняются местами по сравнению со случаем (а); в) точка пересечения изоклин – точка равновесного состояния обоих видов, обладающая локальной устойчивостью. Область 4 – сосуществование двух видов; области 2, 3 – доминирование вида В; область 1 – вымирание обоих видов; г) точка пересечения – точка равновесного состояния, не обладающая локальной устойчивостью. Результаты конкуренции идентичны результатам случая в), за исключением того, что в области 4* верх одерживает либо вид А, либо вид В в зависимости от начальных условий.
Границами между областями 3, 4, 5 являются линии, проходящие через точку равновесного состояния и имеющие крутизну вектора потребления вида А и вида В. Продемонстрировать этот факт можно с помощью векторного уравнения ![]() или, в раскрытом виде
или, в раскрытом виде
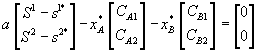 .
.
Для того, чтобы равновесные плотности ![]() были больше 0, необходимо выполнение неравенства
были больше 0, необходимо выполнение неравенства ![]() .
.
Используя описанный метод, результат конкурентной борьбы можно описать для всех возможных классов средств существования.
3.4. Моделирование влияния миграции видов на устойчивость сообщества. Другой аспект применения систем дифференциальных уравнений для описания динамики биологического сообщества – изучение влияния миграции видов на устойчивость сообщества. Д.О.Логофет (1978) исследует следующую схему миграции. Рассмотрены n местообитаний, внутри которых динамика описывается системой
![]()
где xi(t) – численность i-го вида сообщества, в сообществе w видов (w![]() ); функции fi – достаточно гладкие и такие, что существует решение
); функции fi – достаточно гладкие и такие, что существует решение ![]()
![]() и
и ![]() > 0 для всех i.
> 0 для всех i.
Схема миграционного потока задается матрицей ![]() ; элемент матрицы mks указывает поток мигрантов из местообитания s в местообитание k; элемент
; элемент матрицы mks указывает поток мигрантов из местообитания s в местообитание k; элемент ![]() (со знаком “ – ”) дает сумму потоков эмигрантов из k-го местообитания.
(со знаком “ – ”) дает сумму потоков эмигрантов из k-го местообитания.
Для системы, замкнутой по миграции, выполняется соотношение
![]()
Среда предполагается изотропной по миграции, что для матрицы M(n) означает равенство ![]() и равенство единице всех ненулевых недиагональных элементов. Число мигрантов любого вида на любом маршруте в единицу времени считается пропорциональным численности данного вида в том местообитании, откуда происходит эмиграция. Биологические особенности видов описываются параметрами
и равенство единице всех ненулевых недиагональных элементов. Число мигрантов любого вида на любом маршруте в единицу времени считается пропорциональным численности данного вида в том местообитании, откуда происходит эмиграция. Биологические особенности видов описываются параметрами ![]() . Таким образом, динамика композиции сообществ, занимающих n местообитаний, будет описываться уравнениями
. Таким образом, динамика композиции сообществ, занимающих n местообитаний, будет описываться уравнениями
![]()
Исследование состояния равновесия этой системы позволяет сделать вывод о том, что при объединении подсистем с помощью миграции в изотропной среде свойства устойчивости не улучшаются по сравнению с изолированным случаем.
3.5. Управлене ростом и урожаем микроводорослей. Поиск путей оптимизации структуры фитопланктонных сообществ и направленное изменение их структуры имеют как теоретическое (выявление общих механизмов управления экосистемами), так и практическое значение. Изменение качественного и количественного состава сообщества возможно в результате варьирования уровня ресурсов. Экологофизические различия видов в способности использовать тот или иной ресурс ведут к различной конкурентоспособности видов в определенных условиях. Тем самым изменение в доступности какого-либо ресурса будет влиять на результат межвидовой конкуренции (Левич, Личман, 1992).
Вопросы, связанные с управлением ростом и урожаем водорослей в марикультуре, изучались в работе В.А.Силкина и К.М.Хайлова (1988). Рассматривали два основных способа реализации процесса выращивания: периодический и непрерывный. Задача управления в периодической культуре сводится к определению концентрации лимитирующего рост элемента в начальный момент времени для получения необходимого урожая. В непрерывной культуре задача управления формулируется как нахождение оптимальных (по критерию производительности системы) скорости протока D в случае хемостата и плотности биомассы (концентрации клеток) в случае турбидостата.
Для отражения взаимосвязи между удельной скоростью роста и внутриклеточным содержанием элемента питания применяли формулу Друпа
![]()
зависимость скорости поглощения от концентрации элемента питания выражалась уравнением Михаэлиса-Ментен
![]() ,
,
где ![]() – максимальная скорость поглощения; s – концентрация элемента питания; Ks – константа полунасыщения.
– максимальная скорость поглощения; s – концентрация элемента питания; Ks – константа полунасыщения.
Рост водорослей в периодической культуре с элементами минерального питания в качестве лимитирующих факторов описывается уравнениями
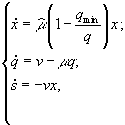
где x – концентрация биомассы клеток; q – внутриклеточное содержание элемента питания; qmin – минимальное содержание биогена в клетке, при котором удельная скорость роста μ равна нулю.
Оптимальное соотношение двух элементов по критерию удельной скорости роста различается в зависимости от стадии роста. В лаг-фазе это соотношение равно
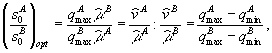
а на стационарной фазе
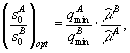
здесь ![]() – максимальные удельные скорости роста культуры при лимитировании веществами A и В, соответственно.
– максимальные удельные скорости роста культуры при лимитировании веществами A и В, соответственно.
При соблюдении условия ![]() оптимальное соотношение элементов не зависит от фазы роста
оптимальное соотношение элементов не зависит от фазы роста
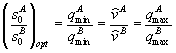 .
.
Поведение популяции одноклеточных организмов при проточном культивировании описывается уравнениями
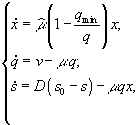
здесь D – скорость протока; s0 – концентрация питательного вещества во входящем потоке; s – концентрация питательного вещества в среде; остальные обозначения сохранены. Критическая скорость разбавления и оптимальная по критерию продукции скорость протока находились для разных значений параметра ![]() и константы полунасыщения Ks:
и константы полунасыщения Ks:
1) ![]() :
: ![]()
2) ![]() :
: ![]()
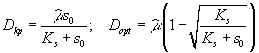
Исследование влияния различных концентраций азота и фосфора в среде на структуру лабораторного водорослевого сообщества, функционирующего в условиях накопительного культивирования, было проведено в работе А.П.Левича и Е.Г.Личман (1992). Согласно модели С.Йоргенсена (1985) для каждого вида ![]() в модель включены три уравнения: динамики биомассы и динамики внутриклеточного содержания азота и фосфора, а для всего сообщества – уравнения динамики концентраций азота и фосфора в среде:
в модель включены три уравнения: динамики биомассы и динамики внутриклеточного содержания азота и фосфора, а для всего сообщества – уравнения динамики концентраций азота и фосфора в среде:
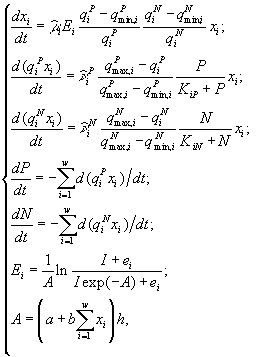
где xi – концентрация биомассы i - го вида; ![]() – клеточная квота в единице биомассы по фосфору;
– клеточная квота в единице биомассы по фосфору; ![]() – клеточная квота в единице биомассы по азоту; N, P –концентрация азота и фосфора в среде;
– клеточная квота в единице биомассы по азоту; N, P –концентрация азота и фосфора в среде; ![]() – максимальная удельная скорость вида;
– максимальная удельная скорость вида; ![]() – максимальные удельные скорости поглощения фосфора и азота i-м видом;
– максимальные удельные скорости поглощения фосфора и азота i-м видом; ![]() – минимальная и макимальная клеточные квоты по фосфору и азоту соответственно;
– минимальная и макимальная клеточные квоты по фосфору и азоту соответственно; ![]() – константы полунасыщения по фосфору и азоту для i-го вида; ei – константа освещенности для i-го вида; I – уровень освещенности; a – коэффициент поглощения света водой; b – коэффициент поглащения света фитопланктоном; h – глубина. В данной модели учтено световое лимитирование: эффект самозатемнения с нарастанием биомассы и разная чувствительность видов к освещенности. Лимитирование биогенными элементами и светом независимы, что отражается присутствием ответственного за световое лимитирование члена в качестве сомножителя.
– константы полунасыщения по фосфору и азоту для i-го вида; ei – константа освещенности для i-го вида; I – уровень освещенности; a – коэффициент поглощения света водой; b – коэффициент поглащения света фитопланктоном; h – глубина. В данной модели учтено световое лимитирование: эффект самозатемнения с нарастанием биомассы и разная чувствительность видов к освещенности. Лимитирование биогенными элементами и светом независимы, что отражается присутствием ответственного за световое лимитирование члена в качестве сомножителя.
Были проведены анализ чувствительности модели, калибровка и верификация. С целью получения необходимых для калибровки и верификации данных был поставлен эксперимент с лабораторными альгоценозами из одного и нескольких видов пресноводных микроводорослей. А именно, два вида зеленых микроводорослей – Scenedesmus. quadricauda и Ankistrodesmus falcatus и цианобактерия Anabaena variabilis выращивали в режиме накопительного культивирования в монокультурах, двух- и трехвидовых поликультурах при различных начальных концентрациях азота и фосфора в среде. При анализе каждой из моделей (модели монокультур, двухвидовых поликультур и трехвидовых) найденные калибровкой параметры верифицировали по данным из других сред. Затем проводили сравнение модельной (с найденными параметрами) и экспериментальной динамик биомасс видов. Во всех случаях наблюдалось хорошее согласие данных. Параллельно с калибровкой и верификацией модели проводили проверку гипотезы о независимости параметров видов от состава сообщества. В результате анализа был сделан вывод о зависимости значений параметров видов от состава сообщества.
С разработанной имитационной моделью были проведены машинные эксперименты по выявлению влияния различных комбинаций азота и фосфора в среде на обилие видов в сообществе. На основе полученных результатов был сделан вывод о существовании реальной возможности управления структурой альгоценозов с помощью варьирования отношений концентраций биогенных элементов, в частности, концентраций азота и фосфора. В ряде случаев возможно добиться увеличения относительного обилия желаемого вида путем создания в среде отношения азота к фосфору, близкого к отношению минимальных квот данного вида. Указания на существование таких оптимальных соотношений имеются в литературе (Smith, 1982; Левич, 1989; см. также п. 4.17). Однако, обнаруженная зависимость не абсолютна (она может проявляться в виде более или менее отчетливых тенденций, а может нивелироваться влиянием различных факторов). Поэтому помимо отношения минимальных квот необходимо учитывать и остальные характеристики видов, которые могут повлиять на конечное распределение относительных обилий видов в сообщеcтве.
4. Экстремальные принципы
4.1. Логистическое уравнение как экстремаль функционала действия. Один из способов применения целевой функции (см. п. 2.2) состоит в формулировании общего утверждения относительно поведения системы. Хорошо известные экстремальные принципы относятся к этому случаю. Самый известный из них – принцип Гамильтона, согласно которому, каждая механическая система ведет себя так, чтобы действие (интеграл по времени от функции Лагранжа) было минимальным. В экологии предпринимались попытки использования этого подхода (Wilhelm, Brüggemann, 2000) для получения уравнения роста популяции, точнее, рассматривалась обратная задача: записать действие, которое приведет к специальному уравнению роста. Одна из наиболее удачных попыток разрешить эту задачу, предложенная М.Гатто с соавторами (Gatto et al., 1988а,b), представлена в работе Дж.Вебба (Webb, 1995).
В качестве функционала действия, который приведет к логистическому уравнению роста популяции численности n, было рассмотрено следующее выражение
Для упрощения вычисления была сделана замена переменных
 (4.1)
(4.1)
Согласно вариационному принципу, уравнение эволюции x(t) задается требованием экстремальности действия, т.е. dS = 0. После необходимых вычислений было получено динамическое уравнение
![]()
![]() (4.2)
(4.2)
Чтобы сравнить этот результат с логистическим уравнением ![]() его переписали в переменных
его переписали в переменных ![]() и продифференцировали:
и продифференцировали: ![]() Полученное совпадение показывает, что любое решение логистического уравнения является решением динамического уравнения, выведенного из функционала действия. Однако, не любое решение уравнения (4.2) является решением логистического уравнения. Для выявления взаимосвязи между данными уравнениями было проведено исследование полученного уравнения эволюции. После некоторых преобразований и интегрирования было получено выражение
Полученное совпадение показывает, что любое решение логистического уравнения является решением динамического уравнения, выведенного из функционала действия. Однако, не любое решение уравнения (4.2) является решением логистического уравнения. Для выявления взаимосвязи между данными уравнениями было проведено исследование полученного уравнения эволюции. После некоторых преобразований и интегрирования было получено выражение
![]()
Уравнение эволюции характеризуется константой R: при R > 0 популяция неограниченно растет, при R < 0 популяция достигает максимального значения, а затем уменьшается до 0. Значение R = 0 приводит к логистическому уравнению, тем самым, показывая, что логистический рост – это особый случай равновесия между неограниченным ростом и затуханием.
В работе также был рассмотрен вопрос об интерпретации введенного таким образом “биологического” действия. Описание в терминах кинетической и потенциальной энергии неприемлемо, поскольку ведет к неизменности общей энергии системы (экологические системы обычно подразумеваются открытыми). По аналогии с физикой, где действие разделено на свободное движение и взаимодействие, предлагалось рассматривать действие (4.1) как сумму члена, описывающего популяцию, которая не подвержена помехам в росте, и члена V(x), описывающего внешнее влияние среды на популяцию. Однако, подобная интерпретация хорошо описывает лишь случай V(x) = 0, когда применение вариационного принципа приводит к уравнению экспоненциального роста. Сам М.Гатто и его соавторы описывали действие как цену роста.
По мнению Дж.Вебба, применение вариационного принципа позволяет сместить акцент с поведения системы на факторы, которые его определяют, а также делает возможным разделение внутреннего поведения популяции и эффектов внешней среды.
4.2. Принцип оптимальной конструкции. Вопрос об оптимальности в биологии подробно изучался Р.Розеном (1969). Основой для его рассуждений является естественный отбор, в результате которого живые организмы приобретают признаки, оптимальные для данных условий, т.е. такие, которые не скажутся отрицательно на их способности конкурировать с соперниками. Основная гипотеза заключается в том, что организмы, обладающие биологической структурой, оптимальной в отношении естественного отбора, оптимальны также и в том смысле, что они минимизируют некоторую оценочную функцию (принцип оптимальной конструкции). Эта функция определяется исходя из основных характеристик окружающей среды.
 Одним из приложений принципа оптимальной конструкции является моделирование кровеносной системы. Рассмотрена задача нахождения оптимального угла отклонения боковой ветви от основного ствола. Радиусы ствола и ответвления считаются известными: r0 и r1. В качестве оценочной функции выбирается сопротивление кровеносной системы, которое согласно принципу оптимальности должно быть минимальным (участок АДС на рис. 16).
Одним из приложений принципа оптимальной конструкции является моделирование кровеносной системы. Рассмотрена задача нахождения оптимального угла отклонения боковой ветви от основного ствола. Радиусы ствола и ответвления считаются известными: r0 и r1. В качестве оценочной функции выбирается сопротивление кровеносной системы, которое согласно принципу оптимальности должно быть минимальным (участок АДС на рис. 16).
Рис.16. Отхождение бокового сосуда от основного ствола. q – угол ответвления
Зависимость полного сопротивления RT участка АДС от угла ответвления q выглядит следующим образом
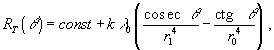
где k – коэффициент пропорциональности, зависящий от вязкости и плотности жидкости; λ0 – длина отрезка СВ.
Дифференцирование функции RT(q ) по q и приравнивание результата нулю дает оптимальное значение угла
![]() .
.
 Следующим шагом является рассмотрение разветвления сосудов (рис. 17).
Следующим шагом является рассмотрение разветвления сосудов (рис. 17).
Рис.17. Разветвление сосудов
Оценочной функцией здесь выступает мощность, рассеиваемая при движении жидкости
![]() ,
,
где f – поток жидкости; K – постоянный коэффициент пропорциональности; RT – полное сопротивление; V – объем изучаемого участка.
Полученные оптимальные значения углов разветвления сравнивали с реальными углами системы кровеносных сосудов кошки и получили хорошие совпадения.
Развитие описанных методов позволяет найти оптимальный радиус отходящей ветви, радиус аорты, а также общее число капилляров, в предположении, что каждый сосуд (кроме аорты) образуется в результате разветвления более крупного сосуда. Полученные результаты хорошо совпадают с эмпирическими данными.
На основании принципа оптимальной конструкции Р.Розеном также была высказана гипотеза о том, что аллометрические соотношения, имеющие место в живой природе, могут быть выведены из условия минимизации некоторого функционала, и показывается, как эта гипотеза может объяснить теорию трансформации д'Арси Томпсона.
4.3. Принцип максимума мальтузианского параметра. Пусть сообщество состоит из w популяций. В каждый момент времени оно может быть описано численностями (или биомассами) составляющих его популяций xi. Пусть ![]() – суммарная биомасса сообщества. Пусть каждая популяция характеризуется мальтузианским параметром
– суммарная биомасса сообщества. Пусть каждая популяция характеризуется мальтузианским параметром ![]() из уравнения
из уравнения ![]() . Пусть
. Пусть ![]() – относительное обилие i-го вида в сообществе. Тогда набор p = {p1,…,pw} – называется структурой сообщества; величина
– относительное обилие i-го вида в сообществе. Тогда набор p = {p1,…,pw} – называется структурой сообщества; величина ![]() – средним мальтузианским параметром сообщества (
– средним мальтузианским параметром сообщества (![]() ), а динамика общей биомассы описывается уравнением
), а динамика общей биомассы описывается уравнением ![]()
Постулируется принцип максимума среднего мальтузианского параметра: сообщество взаимодействующих популяций эволюционирует таким образом, что его средний мальтузианский параметр всегда возрастает, достигая в устойчивом равновесии максимума (Свирежев, 1991).
Применение методов математической статистики и теории вероятностей позволяет изучить вопрос о реализации принципа максимума мальтузианского параметра (в предположении, что постулат справедлив) за счет изменения структуры сообщества. Так, в цитируемой работе определены вектор, задающий структуру сообщества в устойчивом равновесии, и вероятность этого равновесия. Кроме того, возможно разделение структур на вероятные и маловероятные. Получены условия, при которых в процессе адаптации из сообщества эллиминируютия все виды, кроме одного или ни один вид из сообщества не исключается.
4.4. Принцип выживания. В качестве критерия оптимальности предлагается использовать принцип выживания, полагая, что в диаде выживание – приспособленность первичным является выживание (Ханин, 1982).
Пусть динамику экосистемы, в которую входит рассматриваемый вид, адекватно описывает система уравнений с неизвестными численностями особей всех элементов экосистемы. В качестве параметров уравнений выступают экологические условия, а также структурно-функциональные параметры особей всех элементов экосистемы. Выделяют s-я популяция и некоторый структурный или функциональный параметр ![]() этой популяции. Делают предположение о том, что популяция состоит из двух подпопуляций, различающихся величиной фенотипического параметра. Пусть xs(1), xs(2),
этой популяции. Делают предположение о том, что популяция состоит из двух подпопуляций, различающихся величиной фенотипического параметра. Пусть xs(1), xs(2), ![]() ,
,![]() – численности и величины фенотипического параметра двух подпопуляций.
– численности и величины фенотипического параметра двух подпопуляций.
Исследование динамической системы, в которую внесены соответствующие изменения, учитывающие различия фенотипического параметра у особей s-ой популяции, позволяет анализировать асимптотические свойства численностей подпопуляций. Один из возможных вариантов поведения – вытеснение второй подпопуляции первой (фенотипический параметр ![]() имеет селективное преимущество по сравнению с параметром
имеет селективное преимущество по сравнению с параметром ![]() в заданных экологических условиях). Математически этот вариант описывается выражениями
в заданных экологических условиях). Математически этот вариант описывается выражениями
![]()
Оптимальной с точки зрения выживания величиной фенотипического параметра ![]() является такая величина, при которой для любого отличного от этого значения параметра
является такая величина, при которой для любого отличного от этого значения параметра ![]() выполняются условия
выполняются условия
![]()
Причем они верны при произвольных начальных условиях. С оптимальной величиной, удовлетворяющей критерию, следует сопоставлять среднее значение фенотипического параметра.
Необходимо отметить, что если популяция не обладает оптимальным значением параметра, то это не значит, что она элиминируется из биоценоза. Однородная популяция может стабильно существовать при любом значении структурно-функционального параметра ![]() , относящимся к области, соответствующей условию стабильного существования популяции, в частности и при значении, не равном оптимальному
, относящимся к области, соответствующей условию стабильного существования популяции, в частности и при значении, не равном оптимальному![]() . Оптимальное же значение устанавливается в результате конкуренции особей с различными значениями рассматриваемого структурно-фенотипического параметра. Именно вследствие этой конкуренции особи с неоптимальными значениями параметра
. Оптимальное же значение устанавливается в результате конкуренции особей с различными значениями рассматриваемого структурно-фенотипического параметра. Именно вследствие этой конкуренции особи с неоптимальными значениями параметра ![]() элиминируются (Ханини и др., 1978).
элиминируются (Ханини и др., 1978).
Применение общего критерия оптимальности возможно путем численного интегрирования уравнений динамики экосистемы при различных величинах рассматриваемого фенотипического параметра. Также возможно применение частных критериев оптимальности, справедливых в конкретных случаях и следующих из общего критерия. Используя критерий отбора, необходимо учитывать ограничения, вытекающие из физико-химических или биологических закономерностей процесса.
В ряде исследований в качестве критерия оптимальности выступало требование максимума относительной скорости роста численности популяции:
![]()
Этот критерий может быть применен для определения оптимальных величин структурно-функциональных параметров, если относительная скорость роста численности представлена в виде функции этих параметров. Причем, если рассматриваемый параметр не зависит от возраста особи, то задача нахождения оптимального значения сводится к отысканию параметра, соответствующего максимуму относительной скорости роста; если же рассматриваемый параметр зависит от возраста, то искомая оптимальная зависимость может быть определена путем решения соответствующей вариационной задачи (Ханин и др., 1978).
Общий критерий оптимальности применяли к исследованию популяций лосей в лесном биоценозе. Оптимизируемыми параметрами были начальный вес новорожденных и рождаемость. Кроме того, из общего критерия оптимальности выводили требование максимума относительной скорости роста популяции, а затем на основании этого требования оптимизировали функцию роста, определяющая зависимость веса тела особи от возраста. Сравнение теоретических величин, полученных для лосей, и соответствующих биологических данных свидетельствовали об их хорошем согласии.
В теории оптимальных биологических процессов применимы более простые критерии, например, определяющие оптимальность структурно-функциональных параметров органов и систем, роль которых в организме сводится к выполнению определенных функций. Критерием оптимальности такого органа является условие минимума его потребностей при условии выполнения этим органом заданных функций
![]() (4.3)
(4.3)
где Пор – потребности органа; Пп – потребление пищи в единицу времени, связанное с поддержанием жизненного органа, не несущего функциональную нагрузку; Пf – потребление пищи в единицу времени, связанное с осуществлением органом его функций в организме. Использование критерия (4.3) требует учитывать условия, определяющие функции, выполняемые органом или системой.
Критерий, определяющий оптимальные функциональные параметры, имеет вид: Пf = min. Здесь необходимо сформулировать дополнительные условия, определяющие функции органа.
Если определяющей является энергетическая деятельность органа, то критерий оптимальности может быть сформулирован в виде ![]() , где Wi – мощность, потребляемая i-м органом.
, где Wi – мощность, потребляемая i-м органом.
Авторами цитируемой работы представлено применение общего критерия отбора для определения оптимального в эволюционном смысле начального веса новорожденных (на примере данных биологических исследований для популяции лосей); энергетического критерия оптимальности (4.3) для определения функционального состояния системы транспорта кислорода при физической нагрузке и при ее отсутствии, а так же для нахождения энергетически оптимальной концентрации эритроцитов в крови, парциального давления в артериальной и венозной крови, определения оптимальных функциональных параметров системы внешнего дыхания и др.
4.5. Принцип максимума использованной энергии. Н.С.Печуркин (1982) предложил энергетический принцип экстенсивного развития, согласно которому, в процессах биологического развития надорганизменных систем (эволюции, экологических сукцессиях и перестройках) величина использованного биологической потока энергии возрастает, достигая локальных максимальных значений в стационарных состояниях.
Обозначив через Hпад величину потока падающей энергии, т.е. потока энергии, который потенциально может быть использован некоторой биологической системой, через Hисп – ту часть потока, которую система захватывает и использует, через Hнеисп – неиспользованную часть потока (Hпад= Hисп+ Hнеисп), сформулированный энергетический принцип можно записать в виде:
![]() или
или ![]()
Учитывая, что поток энергии, захваченный системой складывается из доли, связанной с запасами энергии в активной биомассе В, и потерь на дыхание системы R (![]() , μ – показатель прироста биомассы), критерий для внутреннего распределения энергии запишется в виде
, μ – показатель прироста биомассы), критерий для внутреннего распределения энергии запишется в виде![]()
Кинетическое обоснование принципа экстенсивного развития было проведено для модели динамики микробных популяций, эволюционирующих в условиях проточного культивирования. Существование отбора показано при введении двух типов селекционных ограничений: постоянной общей организации и постоянного общего потока. Экспериментальной эволюционной системой, соответствующей условию постоянных потоков, является хемостат, в котором развивается генетически неоднородная популяция микроорганизмов. Система уравнений, описывающих эту ситуацию, имеет вид
 (4.4)
(4.4)
где (1 – ki) – доля точных копий в поколении i-й формы; m i – удельная скорость роста i-й формы микроорганизма; di – удельная скорость отмирания i-й формы;
xi – концентрация i-й формы; D – удельная скорость разбавления; s – концентрация лимитирующего субстрата; s0 – концентрация субстрата во входном потоке; Yi – коэффициенты, определяющие потребление субстрата на прирост биомассы; ![]() – константы полунасыщения.
– константы полунасыщения.
Второе уравнение системы (4.4) можно записать на языке потоков энергии: ![]()
В условиях сильного лимитирования (![]() , где
, где ![]() – все виды элиминации исходной формы) для стационарного состояния выполняется соотношение
– все виды элиминации исходной формы) для стационарного состояния выполняется соотношение
![]()
Появление мутанта с повышенной скоростью роста ![]() будет приводить к его накоплению и снижению концентрации лимитирующего субстрата в стационарном состоянии
будет приводить к его накоплению и снижению концентрации лимитирующего субстрата в стационарном состоянии ![]() . Снижение величины
. Снижение величины ![]() будет вызывать накопление мутантов со сниженной константой полунасыщения
будет вызывать накопление мутантов со сниженной константой полунасыщения ![]() , уменьшенной скоростью отмирания dm < di и пониженной частотой мутирования km < ki. При появлении следующего, более активного мутанта будет происходить переход к еще более низкой стационарной величине
, уменьшенной скоростью отмирания dm < di и пониженной частотой мутирования km < ki. При появлении следующего, более активного мутанта будет происходить переход к еще более низкой стационарной величине![]() . Поскольку скорость протока D остается неизменной, будет возрастать степень использования субстрата. В этом можно убедиться, записав стационарное состояние для второго уравнения системы (4.4)
. Поскольку скорость протока D остается неизменной, будет возрастать степень использования субстрата. В этом можно убедиться, записав стационарное состояние для второго уравнения системы (4.4) ![]() на языке потоков энергии
на языке потоков энергии![]() Таким образом, величина Ннеисп снижается и при неизменном значении Нпад увеличивается величина Нисп, т.е. для системы с постоянными потоками выполняется принцип экстенсивного развития.
Таким образом, величина Ннеисп снижается и при неизменном значении Нпад увеличивается величина Нисп, т.е. для системы с постоянными потоками выполняется принцип экстенсивного развития.
Для системы с постоянной организацией (типа турбидостата) динамика описывается уравнениями
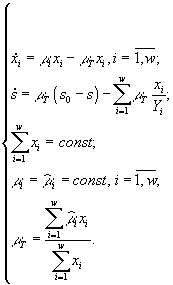
Балансовое уравнение потоков имеет вид ![]() или
или ![]() В результате эволюции системы увеличивается скорость разбавления m
Т при сохранении концентрации биомассы, а, значит, и возрастает поток использованной энергии. Хотя поток неиспользованной энергии тоже будет возрастать (значение
В результате эволюции системы увеличивается скорость разбавления m
Т при сохранении концентрации биомассы, а, значит, и возрастает поток использованной энергии. Хотя поток неиспользованной энергии тоже будет возрастать (значение ![]() не меняется согласно уравнению для стационарного состояния
не меняется согласно уравнению для стационарного состояния ![]() ), его прирост будет компенсирован гораздо большим возрастанием потока падающей энергии.
), его прирост будет компенсирован гораздо большим возрастанием потока падающей энергии.
В той же работе Н.С.Печуркин сформулировал энергетический принцип интенсивного развития: любая живая система надорганизменного уровня развивается (эволюционирует) таким образом, что поток использованной энергии на единицу биологической структуры за время ее существования возрастает.
Показатель уровня интенсивного энергетического развития ![]() может быть записан в виде
может быть записан в виде ![]() . Если за время генерации принять время удвоения биомассы, то
. Если за время генерации принять время удвоения биомассы, то ![]() (g – длительность поколения).
(g – длительность поколения).
Для меняющегося потока 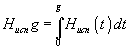 , где Нисп(t) – мгновенное значение потока энергии.
, где Нисп(t) – мгновенное значение потока энергии.
Для демонстрации выполнения сформулированного принципа приводятся следующие примеры:
1) для микроорганизмов ![]() 1,7 – 2,3;
1,7 – 2,3;
для растений ![]() 1,82;
1,82;
для насекомых ![]() 7 – 10;
7 – 10;
для млекопитающих ![]() 50 – 100;
50 – 100;
2) зависимость интенсивности теплорассеяния Q от веса организма M описывается уравнением: ![]() , a, k – константы. Коэффициент a, характеризующий интенсивность энергообмена, в процессе эволюции существенно возрастает: для одноклеточных а = 0,084 кал/ч, для пойкилотермных животных а = 0,69 кал/ч, для гомойтермных а = 19,68 кал/ч.
, a, k – константы. Коэффициент a, характеризующий интенсивность энергообмена, в процессе эволюции существенно возрастает: для одноклеточных а = 0,084 кал/ч, для пойкилотермных животных а = 0,69 кал/ч, для гомойтермных а = 19,68 кал/ч.
Для конкретных приложений (например, к классификации в пределах вида или рода) применимость энергетического принципа интенсивного развития как универсального критерия макроэволюции пока разработана недостаточно.
4.6. Оптимальная жизненная стратегия распределения энергетических ресурсов индивида. Методы вариационного исчисления применяются в эволюционной экологии. Оптимизационный подход основан на предположении, что в процессе эволюции вида при заданных внешних условиях и физиологических ограничениях оптимизируется удельная скорость роста популяции, которая выступает в качестве критерия дарвиновской приспособленности. Это позволяет формулировать задачу эволюционной оптимизации жизненного цикла в терминах математической теории оптимального управления как задачу нахождения оптимальных жизненных стратегий распределения энергетических ресурсов индивида между ростом, размножением, добыванием пищи, защитой от неблагоприятных влияний внешней среды и репарацией (вклад энергии в репарацию означает вклад в “будущее выживание”) (Терехин, 2001).
Общая оптимизационная задача выглядит следующим образом (Teriokhin, 1998)

Здесь u(t) – часть энергии, затраченная на воспроизведение; l(t) – функция выживания; d(t) – смертность, включающая четыре компоненты: две контролируемые и две неконтролируемые индивидом; a – постоянная часть неконтролируемой (внешней) смертности; bt – неконтролируемая смертность, возрастающая с возрастом со скоростью b; p(t) – текущий уровень смертности, регулируемый вкладом энергии v(t) – (часть энергии направленная на выживание); c ³ 0, c = const; q(t) – скорость старения, контролируемая вкладом энергии в репарацию w(t). Последнее равенство означает, что никакие другие нужды организма, кроме воспроизводства, выживания и репарации, не включаются в модель.
Целевым функционалом является жизненный репродуктивный успех особи. Этот функционал применим в тех случаях, когда численность популяции меняется не слишком быстро, иначе, правильнее было бы использовать удельную скорость роста численности популяции μ, получаемую из уравнения Эйлера-Лотки ![]() (Терехин, 2001). Однако, рассматривая множитель
(Терехин, 2001). Однако, рассматривая множитель ![]() как часть выживания, можно считать, что оптимизация удельной скорости μ эквивалентна оптимизации R0 в присутствии дополнительной константы внешней смертности μ (Teriokhin, 1998).
как часть выживания, можно считать, что оптимизация удельной скорости μ эквивалентна оптимизации R0 в присутствии дополнительной константы внешней смертности μ (Teriokhin, 1998).
Изучение модели идет по пути рассмотрения частных случаев. Задача о распределении энергии между воспроизведением и выживанием описывается исходной системой при условии, что w(t) º
0. Вариационная задача решается применением принципа максимума Понтрягина. Оптимальная стратегия задается выражением: ![]() , где функция x(t) – решение уравнения
, где функция x(t) – решение уравнения ![]()
Численные решения этих уравнений позволяют наблюдать зависимость стратегии от параметров.
Предположение о неопределенности максимальной продолжительности жизни Т (Т = ¥
) и постоянстве u(t) º
U дает оптимальное значение U = 0.5 (при a = b = d = 0); 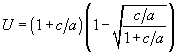 (при b = d = 0).
(при b = d = 0).
Условие v(t) º 0 означает распределение затрат энергии между воспроизводством и репарацией. В этом случае гамильтониан выглядит следующим образом
![]() где
где
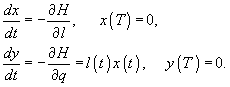
Для нахождения оптимальной стратегии необходимо решить четыре дифференциальных уравнения, что можно сделать только численными методами. Другой путь – дискретизация задачи и применение динамического программирования, последнее позволяет построить графики зависимости u от t и q, q от t для разных наборов параметров a, b, c.
Важным результатом решения этой частной задачи является получение увеличения скорости смертности с возрастом. Данная зависимость хорошо согласуется (по крайней мере качественно) с реальными наблюдениями. Обычно ускоренный рост интенсивности смертности с возрастом описывается уравнением Гомпертца-Макехама ![]() (α, β, γ – неотрицательные константы; t – возраст) или уравнением Вейбулла (показательный рост). Получение же такой зависимости из вариационной задачи позволяет объяснить ее требованием эволюционной оптимальности, а именно, необходимостью увеличения с возрастом трат энергии на размножение за счет уменьшения трат на репарацию (Teriokhin, 1998; Терехин, 2001).
(α, β, γ – неотрицательные константы; t – возраст) или уравнением Вейбулла (показательный рост). Получение же такой зависимости из вариационной задачи позволяет объяснить ее требованием эволюционной оптимальности, а именно, необходимостью увеличения с возрастом трат энергии на размножение за счет уменьшения трат на репарацию (Teriokhin, 1998; Терехин, 2001).
Задача о распределении энергии между выживанием и репарацией также решается с помощью динамического программирования.
Другой аспект изучения распределения организмом энергетических затрат на рост и размножение заключается в попытке объяснить общую закономерность развития многих животных и растений, которая состоит в том, что размножение начинается тогда, когда рост в основном закончен (Инсаров, 1975). Предлагается следующий экстремальный принцип: в онтогенезе комбинация между тратами на рост и размножение оптимальная в отношении естественного отбора организмов такова, что биомасса потомства одной особи за все время ее жизни достигает максимума. Для формализации этого принципа вводят обозначения: M(t) – масса ассимиляционного аппарата организма в зависимости от времени t (0 £ t £ T); j (t) – доля M(t), идущая на рост самого организма, 0 £ j (t) £ 1; y (t) – средняя вероятность того, что организм доживет до момента времени t, y (0) = 1, y (Т) = 0.
Предполагается, что за время жизни организм не расходует все свои ресурсы только на рост (![]() ), и все, что не тратится на рост, идет на размножение (1 – j
(t) – доля M(t), расходуемая на размножение). Функция ¦
(t) считается известной и является коэффициентом пропорциональности между приростом M(t) в константной среде и j
(t)×
M(t) (dM(t) = ¦
(t)×
j
(t)×
M(t)dt).
), и все, что не тратится на рост, идет на размножение (1 – j
(t) – доля M(t), расходуемая на размножение). Функция ¦
(t) считается известной и является коэффициентом пропорциональности между приростом M(t) в константной среде и j
(t)×
M(t) (dM(t) = ¦
(t)×
j
(t)×
M(t)dt).
Учитывая принятые обозначения, можно записать выражение для массы ассимилятов, потраченной средним организмом на размножение
 .
.
Итак, задача состоит в следующем: найти такую функцию j
0(t), что ![]() , причем множество {j
} – множество кусочно-непрерывных функций j
(t) – таких, что 0 £
j
(t) £
1 для всех t. Предположение о том, что организм, “управляемый” функцией j
0(t), обладает максимальной биомассой потомства делается на основе тесной связи величины F(j
) и биомассы потомства.
, причем множество {j
} – множество кусочно-непрерывных функций j
(t) – таких, что 0 £
j
(t) £
1 для всех t. Предположение о том, что организм, “управляемый” функцией j
0(t), обладает максимальной биомассой потомства делается на основе тесной связи величины F(j
) и биомассы потомства.
Решение j 0(t) является ступенчатой функцией, принимающей значения 0 и 1, причем, если j 0(t) = 0 при 0 £ t £ b1 (b1 – первая точка переключения функции j 0(t)), то такая функция j 0(t) есть “постороннее” решение, которому не соответствует никакой организм (иначе первое накопление биомассы наступило бы после размножения). Биологический смысл указанного решения заключается в том, что в каждый момент времени организм тратит все ресурсы либо на рост, либо на размножение.
По мнению авторов модели, наиболее адекватно с помощью принципа оптимальности может быть описан рост и размножение некоторых растений, в частности однолетних деревьев, некоторых ракообразных и теплокровных животных, а различные несоответствия в описании развития организмов могут быть объяснены неучтенностью влияния изменения внешней среды, инерцией роста и другими причинами.
Основной результат заключается в том, что тенденция резкого разделения во времени роста и размножения выведена из принципа оптимальности.
4.7. Экстремальный принцип в описании микробиологических процессов. Математическая модель, основанная на вариационных принципах была использована для описания явления диауксии (рост бактерий на смеси субстратов разной доступности), а также для нахождения макроскопического показателя выхода биомассы по стехиометрии индивидуальных метаболических реакций (Паников, 1991). Рассмотрена следующая схема. Рост микроорганизмов представлен как результат совокупности метаболических реакций, в результате которых экзосубстраты превращаются в клеточные компоненты. Каждый из бесконечного множества внутриклеточных путей имеет свою “цену” в виде расхода лимитирующего вещества на построение единицы биомассы, но существует единственный маршрут с наименьшей ценой. В случае хемостата принято, что цена ![]() стремится к минимуму. При заданных скорости потока D и начальной концентрации субстрата s0 минимальному значению цены соответствует максимальная продуктивность хемостата
стремится к минимуму. При заданных скорости потока D и начальной концентрации субстрата s0 минимальному значению цены соответствует максимальная продуктивность хемостата ![]() (μ – удельная скорость прироста биомассы). В соответствии с предлагающимся экстремальным принципом, клеткой выбирается именно этот маршрут с помощью естественных регуляторных механизмов. В качестве примера рассмотрена модель роста, лимитированного источником азота. Выделено 6 переменных – одна внешняя и пять внутренних: интередиаты L, а также высокомолекулярные соединения, которые синтезируются из интередиатов L – транспортные белки Р1, ферменты небелкового синтеза Р2, рибосомальная РНК R, и прочие каталитически неактивные компоненты биомассы
(μ – удельная скорость прироста биомассы). В соответствии с предлагающимся экстремальным принципом, клеткой выбирается именно этот маршрут с помощью естественных регуляторных механизмов. В качестве примера рассмотрена модель роста, лимитированного источником азота. Выделено 6 переменных – одна внешняя и пять внутренних: интередиаты L, а также высокомолекулярные соединения, которые синтезируются из интередиатов L – транспортные белки Р1, ферменты небелкового синтеза Р2, рибосомальная РНК R, и прочие каталитически неактивные компоненты биомассы ![]() . Пул низкомолекулярных N-соединений L непрерывно восполняется за счет активного транспорта и расходуется на биосинтез четырех макромолекулярных компонентов по разветвленной цепочке. В первой точке ветвления разделение потока интередиатов L осуществляется по соотношению внутреклеточных концентраций рибосомальных РНК R и ферментов Р2. Далее каждый из потоков делится еще раз специальными регуляторными операторами a
(распределение потока L между белками Р1 и ферментами Р2) и b
(распределение потока L между концентрациями рибосомальных РНК R и биомассой
. Пул низкомолекулярных N-соединений L непрерывно восполняется за счет активного транспорта и расходуется на биосинтез четырех макромолекулярных компонентов по разветвленной цепочке. В первой точке ветвления разделение потока интередиатов L осуществляется по соотношению внутреклеточных концентраций рибосомальных РНК R и ферментов Р2. Далее каждый из потоков делится еще раз специальными регуляторными операторами a
(распределение потока L между белками Р1 и ферментами Р2) и b
(распределение потока L между концентрациями рибосомальных РНК R и биомассой ![]() ), a
, b
Î
[0, 1]. Каждой паре операторов a
, b
отвечает один единственный набор установившихся концентраций всех выделенных переменных. Таким образом, операторы a
и b
могут быть использованы как инструменты оптимального управления. Модель описывается системой уравнений
), a
, b
Î
[0, 1]. Каждой паре операторов a
, b
отвечает один единственный набор установившихся концентраций всех выделенных переменных. Таким образом, операторы a
и b
могут быть использованы как инструменты оптимального управления. Модель описывается системой уравнений
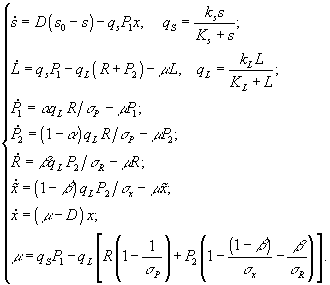
Здесь Ks, KL – константы полунасыщения; qs, qL – удельные скорости потребления ресурса; ks, kL – максимальные удельные скорости потребления ресурса; σx, σP – клеточные квоты; параметры s0, Ks, KL, ks, kL, σx, σP – константы.
Для нахождения установившегося состояния в хемостате производные в системе уравнений приравниваются нулю, затем решается получившаяся система алгебраических уравнений. Суть оптимизационной задачи состоит в нахождении таких операторов a
и b
, при которых минимизируется расход субстрата на синтез единицы биомассы ![]() во всей области допустимого варьирования скорости протока D (
во всей области допустимого варьирования скорости протока D (![]() – значение биомассы в установившемся режиме). Эта задача эквивалентна поиску максимума биомассы
– значение биомассы в установившемся режиме). Эта задача эквивалентна поиску максимума биомассы ![]() для заданных значений D и s0.
для заданных значений D и s0.
4.8. Принцип максимальной неожиданности протекания эволюции. Один из путей исследования динамики популяций – изучение “дарвиновских систем”, описывающих динамику естественного отбора в биологических популяциях. Одним из исследователей этих объектов является Е.В.Евдокимов, в работе которого приведено определение и способ описания дарвиновских систем по М.Эйгену (Евдокимов, 1999). Дарвиновские системы (ДС) являются открытыми системами, состоящими из самокопирующихся с небольшим количеством ошибок единиц различных видов, использующих для своего размножения вещество и свободную энергию поступающих извне питательных компонентов. Обязательными ограничениями являются либо постоянство суммарной организации системы, либо постоянство потоков. Для описания ДС используют дифференциальное уравнение
![]() , (4.5)
, (4.5)
которое можно свести к достаточно простому виду
![]() (4.6)
(4.6)
где i, j=1,...,w (w = const) – число квазивидов в системе, s = (s1,..., sm) – концентрации питательных компонентов, ![]() – удельная скорость размножения i-го квазивида, D – скорость протока в системе. В зависимости от наложенных ограничений различают ДС с постоянной организацией (ДСПО) – у которых сумма
– удельная скорость размножения i-го квазивида, D – скорость протока в системе. В зависимости от наложенных ограничений различают ДС с постоянной организацией (ДСПО) – у которых сумма ![]() и концентрация s постоянны, и ДС с постоянным протоком (ДСПП), характеризующиеся условием D = const.
и концентрация s постоянны, и ДС с постоянным протоком (ДСПП), характеризующиеся условием D = const.
Изучению ДС посвящено много работ, однако, при экспериментальных исследованиях возникает проблема неполноты их описания с помощью систем (4.5 и 4.6) из-за практической труднодоступности информации на микроуровне (Евдокимов, 1999). Для избежания подобных трудностей была предпринята попытка использовать формализм Джейнса (см. также п.4.17). В своей работе Е.В.Евдокимов предложил модификацию этого метода: “В соответствии с общим подходом потребуем, чтобы процесс эволюции ДС протекал наименее неожиданным способом” (принцип минимальной неожиданности протекания эволюции). В качестве целевой функции используется функция неожиданности эволюции ДСПО
![]()
где ![]() – вероятность того, что случайно выбранная в момент t особь имеет мальтузианский параметр
– вероятность того, что случайно выбранная в момент t особь имеет мальтузианский параметр ![]()
![]() , значения yk0 – задаются экспериментально.
, значения yk0 – задаются экспериментально.
Итак, вариационная задача формулируется следующим образом

Решение было получено методом неопределенных множителей Лагранжа

причем оно соответствует решению системы уравнений (4.6).
Важное значение имеет множитель Лагранжа ![]() , поскольку, придавая ему информационный смысл, можно получить “основную теорему естественного отбора” Фишера
, поскольку, придавая ему информационный смысл, можно получить “основную теорему естественного отбора” Фишера
![]()
Другое важное следствие состоит в том, что множитель λ0 с точностью до коэффициента пропорциональности равен среднему по популяции числу поколений.
При рассмотрении энергетических трат на эволюцию было получено, что множитель λ0 пропорционален энергопотреблению популяции.
Таким образом, исходя из эвристического принципа минимальной неожиданности протекания эволюционного процесса, Е.В.Евдокимов получил результаты, “описывающие динамику отбора в дарвиновских системах, которые полностью идентичны уравнениям, выводимым из кинетики размножения и конкуренции саморедуплицируемых единиц на микроуровне”, а множители Лагранжа, использующиеся для решения вариационной задачи, имеют биологический смысл и прогностическую ценность.
4.9. Модели динамической структуры. В последнее время широкое распространение получило использование так называемой модели динамической структуры. Необходимость применения такого рода моделей обоснована в работах С.Йоргенсена с соавторами (Jørgensen et al., 1995; Bendoriccho, Jørgensen, 1997). Экосистемы и их живые компоненты способны изменять свои свойства с изменением внешних условий, причем регулирующие процессы могут проходить на индивидуальном (адаптация) и на экосистемном (изменение видов и структуры) уровне. Поэтому для описания взаимосвязи между состоянием системы и внешними по отношению к ней факторами необходимо заложить в используемые модели соответствующие экологические законы, позволяющие учесть изменения в составе и (или) структуре системы.
Модель динамической структуры определяется как математическая модель, в рамках которой возможно изменение величины параметров, в соответствии с выбранной целевой функцией (Bendoriccho, Jørgensen, 1997). В отличие от традиционных моделей, в которых имитация действительной ситуации, описываемой экспериментальными данными, идет за счет минимизации суммы разниц между вычисленными и экспериментальными значениями переменных состояния, модели динамической структуры подбираются под реальную картину в результате поиска наилучшей комбинации параметров, которая максимизирует целевую функцию. Ключевой момент такой модели – выбор набора параметров среди большого количества, использующегося в традиционных моделях, и выбор целевой функции, которая наилучшим образом описывает развитие экосистемы. Описание целевых функций, которые в разное время предлагались для применения в экстремальных моделях, приведено в ряде работ (Patten, 1995; Jørgensen et al., 1995; Bendoriccho, Jørgensen, 1997), по которым ведется дальнейшее цитирование.
Общий поток энергии через систему (Total system throughflow of energy) – первая из характеристик системы, предложенная в качестве целевой функции – задается выражением 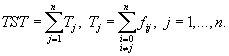 Tj представляет собой общий поток энергии (вещества) из j-ой части системы, fij – поток от j к i, значение индекса i = 0 обозначает внешнюю среду. Согласно принципу максимума мощности, который был развит Х.Одумом и Р.Пинкертоном (Odum, Pinkerton, 1955), функционирование экосистемы происходит так, чтобы максимизировать мощность, т.е., говоря математическим языком, в данной модели требуется максимизировать скорость изменения величины TST. (В случае дискретного времени рассматриваются члены
Tj представляет собой общий поток энергии (вещества) из j-ой части системы, fij – поток от j к i, значение индекса i = 0 обозначает внешнюю среду. Согласно принципу максимума мощности, который был развит Х.Одумом и Р.Пинкертоном (Odum, Pinkerton, 1955), функционирование экосистемы происходит так, чтобы максимизировать мощность, т.е., говоря математическим языком, в данной модели требуется максимизировать скорость изменения величины TST. (В случае дискретного времени рассматриваются члены ![]() где
где ![]() – временной интервал.)
– временной интервал.)
Во всеобъемлющей теории роста и развития экосистемы, предложенной Р.Улановичем (Ulanowicz, 1986), роль целевой функции выполняет асценденция  . Максимизация этой функции должна описывать автокаталитические тенденции, присущие организации развития экологических сетей.
. Максимизация этой функции должна описывать автокаталитические тенденции, присущие организации развития экологических сетей.
Эмержентность была предложена на роль целевой функции Х.Одумом (Odum, 1983). Это качественный фактор энергии, измеряющий удаленность данного типа энергии от первоначальной солнечной энергии. Например, энергия топлива более высокого качества, чем солнечная энергия. 104 калорий солнечной энергии производят 102 калорий первичных продуцентов, которые, в свою очередь, производят 10 калорий хищников. Если принять качество солнечной энергии за единицу, то качество более высоких трофических уровней соответственно будет 100, 1000, 10000.
В последнее время наиболее широкое распространение получила функция эксергии или энергии перехода (Ex), которую понимают как максимальную свободную энергию, которую система способна выделить в пространство при переходе в состояние термодинамического равновесия (Jørgensen, Mejer, 1982). Эксергия определяется окружающей систему средой и может быть рассмотрена как мера термодинамического порядка. Кроме того, энергия перехода непосредственно связана термодинамической информацией I: Ex = IT (T – температура). Для систем с неорганическими потоками и пассивным продуцированием органических веществ энергия перехода задается выражением  где R – газовая постоянная; T – абсолютная температура; ci - концентрация в экосистеме i-го компонента; cieq – соответствующая концентрация i-го компонента в термодинамическом равновесии; индекс i = 0 отвечает неорганическим компонентам рассматриваемого химического вещества. С.Йоргенсен и Х.Мэйер предлагают следующую формулировку термодинамического закона экологии: экосистема стремится ответить на изменения внешних факторов путем формирования нового состава (а может быть даже и новой структуры), максимизирующего при определяемых внешними факторами условиях величину энергии перехода (Jørgensen, Mejer, 1982).
где R – газовая постоянная; T – абсолютная температура; ci - концентрация в экосистеме i-го компонента; cieq – соответствующая концентрация i-го компонента в термодинамическом равновесии; индекс i = 0 отвечает неорганическим компонентам рассматриваемого химического вещества. С.Йоргенсен и Х.Мэйер предлагают следующую формулировку термодинамического закона экологии: экосистема стремится ответить на изменения внешних факторов путем формирования нового состава (а может быть даже и новой структуры), максимизирующего при определяемых внешними факторами условиях величину энергии перехода (Jørgensen, Mejer, 1982).
К.Паттен (Patten, 1995) предложил экстремальный принцип, согласно которому процессы самоорганизации в экосистеме происходит в направлении увеличения косвенных эффектов.
Все перечисленные целевые функции описываются в терминах энергии и сложности экосистемы, что натолкнуло на мысль об их тесной взаимосвязи. В частности, К.Паттен в своей работе (Patten, 1995) показал, что эксергия, эмержентность, сила, асценденция и косвенные эффекты имеют взаимосвязанную микроскопическую динамику.
В процессе упорядоченного развития сообщества экосистема достигает зрелой и стабильной стадии. Стабильность – свойство реагировать на внешние воздействия неболишим изменением биомассы и быстро возвращаться к исходному состоянию после окончания воздействия (Pérez-España, Arreguin-Sánchez, 1999).
Авторы статьи предложили индекс зрелости, основанный на мере неопределенности. Неопределенность понимается как невозможность заранее знать каким будет путь данной единицы вещества или энергии в экосистеме. Было сделано предположение, что стратегия жизни фокусируется в направлении получения как можно более сложной и разнообразной структуры. По этой причине при достижении зрелости экосистема получается более сложной. Сложная система cсоответствует более высокому разнообразию и большому количеству взаимосвязей, что делает доступным большое количество путей для получения энергии в случае каких-либо изменений. В сложной системе уменьшается возможность определить путь данной единицы вещества, и неопределенность возрастает. Формула, выражающая меру неопределенности H для нескольких независимых событий, была описана Р.Улановичем (Ulanowicz, 1986)
![]() ,
,
где pi – вероятность исхода i, n – количество событий, K – константа пропорциональности.
Кроме неопределенности, зрелость должна быть связана с другим параметром, дающим экосистеме независимость от внешних изменений. Авторы полагают, что таким свойством является способность системы использовать ее собственный детрит. Более эффективный цикл использования детрита в системе дает более зрелую, независимую и устойчивую систему. Взяв за основу формулу, выражающую меру неопределенности и добавив потоки детрита, выводится индекс зрелости
 ,
,
где tij – поток вещества от вида i к виду j; T – сумма всех членов tij; ![]() – поток детрита к k-му виду. Чтобы представить все возможные потоки добавляется импорт (индекс 0, только выход), экспорт и дыхание (n + 1 и n + 2, только вход).
– поток детрита к k-му виду. Чтобы представить все возможные потоки добавляется импорт (индекс 0, только выход), экспорт и дыхание (n + 1 и n + 2, только вход).
4.10. Принцип максимизации репродуктивных усилий. В качестве общего критерия экологического моделирования предлагается использовать максимизирование репродуктивных усилий (Zeide, 1991). Рост растительности и все другие процессы должны быть представлены в модели как путь к максимизации усилий направленных на воспроизводство. Вместо простого описания роста или смерти, любой процесс, происходящий в лесу, должен рассматриваться как вклад в конечную цель жизни дерева. Акцентирование внимания на максимизации репродуктивных усилий не обязательно усложняет модель, поскольку для их определения не требуется подсчета каждого семени. Затраты на воспроизведение потомства эквивалентны разнице между усилиями, затраченными на рост и отдачей в форме ассимиляции. Параметры модели подбираются так, чтобы максимизировать эту разницу.
Результат такого подхода часто оказывается неожиданным. Так, например, рост обычно изображается гладкой кривой, дифференцируемой в каждой точке. Эта гладкость скрывает излом, являющийся индикатором начала репродукции. Как было доказано математически, в предсказуемой среде максимум репродуктивных усилий достигается только с полным переключением от вегетативного роста к производству семян. Недостаточная предсказуемость среды сглаживает переход.
Когда рост рассматривается как путь к максимизации репродуктивных усилий, кривая роста может обеспечить гораздо больше информации, в частности, она может обнаружить интенсивность конкуренции, ответственной за время переключения, ожидаемое время жизни и степень предсказуемости среды.
4.11. Дифференциальные уравнения и принцип максимума Понтрягина в биоэкономической модели. Рассматривается простая модель вылова двух конкурирующих видов рыб, подчиняющихся закону логистического роста (Chaudhuri, 1986). Система уравнений, описывающая данную ситуацию, записывается следующим образом

Здесь r, s представляют собой экологический потенциал, K, L – пропускные способности двух видов, члены ![]() и
и ![]() отражают борьбу за общий ресурс, q1 и q2 – коэффициенты вылова видов и E – усилия, затраченные на их совместный вылов. Предполагается, что существует внешний ресурс, поддерживающий каждый вид.
отражают борьбу за общий ресурс, q1 и q2 – коэффициенты вылова видов и E – усилия, затраченные на их совместный вылов. Предполагается, что существует внешний ресурс, поддерживающий каждый вид.
В работе исследуют состояния экологического равновесия, динамику поведения, а также оптимальную стратегию вылова. Чистый доход в любой момент времени определяется формулой: ![]() где С – цена рыбной ловли, пропорциональная усилиям. Предлагается максимизировать общий дисконтированный доход для
где С – цена рыбной ловли, пропорциональная усилиям. Предлагается максимизировать общий дисконтированный доход для ![]() :
: ![]() , δ – мгновенная годовая скорость дисконтирования. Применяя принцип максимума Понтрягина, максимизируется функционал:
, δ – мгновенная годовая скорость дисконтирования. Применяя принцип максимума Понтрягина, максимизируется функционал: ![]() . Были найдены оптимальные равновесные величины популяций, которые ведут к ситуации, когда общая цена улова на единицу усилий равна дисконтированной величине будущей цены. Согласно полученным результатам, при бесконечно большом значении мгновенной годовой скорости дисконтирования δ рыбный промысел должен быть закрыт ввиду полного рассеяния приносимого им дохода, а значение δ = 0 ведет к максимуму дохода, непрерывно получаемого от вылова. Однако, существование такой биоэкономической стабильности зависит от трофической интенсивности обоих видов и неизбежно влечет за собой самоограничение видов.
. Были найдены оптимальные равновесные величины популяций, которые ведут к ситуации, когда общая цена улова на единицу усилий равна дисконтированной величине будущей цены. Согласно полученным результатам, при бесконечно большом значении мгновенной годовой скорости дисконтирования δ рыбный промысел должен быть закрыт ввиду полного рассеяния приносимого им дохода, а значение δ = 0 ведет к максимуму дохода, непрерывно получаемого от вылова. Однако, существование такой биоэкономической стабильности зависит от трофической интенсивности обоих видов и неизбежно влечет за собой самоограничение видов.
4.12. Экстремальные свойства сообщества с горизонтальной структурой. В данном разделе описан несколько иной взгляд на применение экстремальных принципов. Заключается он в получении из динамических уравнений соотношений, которые можно интерпретировать как экстремальные принципы (Свирежев, Логофет, 1978). В основе такого подхода лежит исследование так называемых вольтерровских моделей сообществ, описываемых системой уравнений
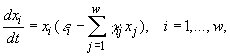
где ![]() – скорость естественного прироста или смертности i-го вида в отсутствие всех остальных видов, а знак и абсолютная величина
– скорость естественного прироста или смертности i-го вида в отсутствие всех остальных видов, а знак и абсолютная величина ![]() отражает соответственно характер и интенсивность влияния j-го вида на i-й вид,
отражает соответственно характер и интенсивность влияния j-го вида на i-й вид, ![]() – показатель внутривидового взаимодействия для i-го вида. В терминах частот (относительных численностей)
– показатель внутривидового взаимодействия для i-го вида. В терминах частот (относительных численностей) ![]()
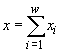 уравнения динамики сообщества имеют вид
уравнения динамики сообщества имеют вид

где 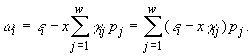 Рассматривается сообщество с горизонтальной структурой, т.е. сообщество, состоящее из видов, расположенных на одном трофическом уровне и конкурирующих за один или несколько ресурсов, что означает выполнение равенств
Рассматривается сообщество с горизонтальной структурой, т.е. сообщество, состоящее из видов, расположенных на одном трофическом уровне и конкурирующих за один или несколько ресурсов, что означает выполнение равенств ![]() .
.
Вектор ![]() с координатами
с координатами ![]() ,
, ![]() называют композицией сообщества. Множество возможных композиций – симплекс
называют композицией сообщества. Множество возможных композиций – симплекс ![]() в положительном ортанте n-мерного пространства. Вектор
в положительном ортанте n-мерного пространства. Вектор ![]() – равновесная композиция, определяемая из условия
– равновесная композиция, определяемая из условия ![]() Показано, что между численностью x и равновесной композицией существует взаимнооднозначное соответствие.
Показано, что между численностью x и равновесной композицией существует взаимнооднозначное соответствие.
Представление уравнений вольтерровской модели в частотной форме позволяет выделить в общем процессе эволюции сообщества два да известной степени различных, хотя и связанных процесса: эволюция композиции и эволюция общей численности. Естественно, что скорости этих процессов могут быть различны.
Вводится функция 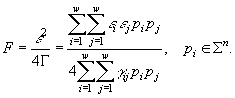 Здесь ε – средний коэффициент естественного прироста для всего сообщества в целом, Г – средний коэффициент конкуренции. Исследование экстремумов функции F дает следующий результат: равновесная композиция
Здесь ε – средний коэффициент естественного прироста для всего сообщества в целом, Г – средний коэффициент конкуренции. Исследование экстремумов функции F дает следующий результат: равновесная композиция ![]() является экстремальной точкой и
является экстремальной точкой и ![]() Функция F может быть интерпретирована как мощность сообщества или мера скорости протекания энергии. Таким образом, формулируется экстремальный принцип: сообщество видов, конкурирующих за один или несколько ресурсов, эволюционирует в сторону увеличения мощности или скорости протекания через него энергии, причем в равновесии эта мощность (или скорость) максимальна.
Функция F может быть интерпретирована как мощность сообщества или мера скорости протекания энергии. Таким образом, формулируется экстремальный принцип: сообщество видов, конкурирующих за один или несколько ресурсов, эволюционирует в сторону увеличения мощности или скорости протекания через него энергии, причем в равновесии эта мощность (или скорость) максимальна.
Однако, этот принцип не говорит о характере (монотонности) эволюции и ее скорости, он является локальным.
Исследование функции 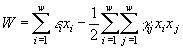 демонстрирует существование глобального максимума и единственного устойчивого состояния равновесия системы
демонстрирует существование глобального максимума и единственного устойчивого состояния равновесия системы 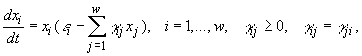 причем любая, начинающаяся внутри положительного ортанта, траектория системы при
причем любая, начинающаяся внутри положительного ортанта, траектория системы при ![]() стремится к данному равновесию. В том случае, когда максимум функции W не достигается внутри положительного ортанта, он будет достигаться на его границе на соответствующих координатных гиперплоскостях. Это означает, что в процессе эволюции экосистемы один или несколько видов должны элиминироваться, поскольку такое равновесное состояние снова будет устойчивым.
стремится к данному равновесию. В том случае, когда максимум функции W не достигается внутри положительного ортанта, он будет достигаться на его границе на соответствующих координатных гиперплоскостях. Это означает, что в процессе эволюции экосистемы один или несколько видов должны элиминироваться, поскольку такое равновесное состояние снова будет устойчивым.
Функция W может быть интерпретирована как разница между репродуктивным потенциалом сообщества ![]() и общими затратами на конкуренцию (на дыхание экосистемы)
и общими затратами на конкуренцию (на дыхание экосистемы) 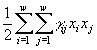 . Тогда возрастание функции W означает стремление системы максимизировать указанную разницу, что можно сделать либо максимизируя репродуктивный потенциал при заданных тратах на конкуренцию, либо минимизируя затраты на конкуренцию при ограниченном репродуктивном потенциале (существуют и промежуточные ситуации). В природе этим двум противоположным механизмам соответствуют, так называемые, r-конкуренция и K - конкуренция.
. Тогда возрастание функции W означает стремление системы максимизировать указанную разницу, что можно сделать либо максимизируя репродуктивный потенциал при заданных тратах на конкуренцию, либо минимизируя затраты на конкуренцию при ограниченном репродуктивном потенциале (существуют и промежуточные ситуации). В природе этим двум противоположным механизмам соответствуют, так называемые, r-конкуренция и K - конкуренция.
Привлечение понятия жизненного пространства, в которое включены все важные для особей сообщества факторы среды, и исследование соответствующих функций, формализующих данное понятие, позволило сформулировать следующий принцип. Сообщество конкурирующих за жизненное пространство видов эволюционирует к состоянию с максимально плотной упаковкой, причем в процессе эволюции плотность упаковки сообщества всегда возрастает, достигая в равновесном состоянии максимально возможного для данной среды значения. (В качестве меры плотности упаковки рассматривается среднеквадратическая разность между реально существующим и необходимым для сообщества жизненным пространством. Чем меньше эта разность, тем плотнее упакованы виды.)
Исследование вольтерровских систем общего вида, т.е. где ![]() могут иметь любой знак, а на коэффициенты
могут иметь любой знак, а на коэффициенты ![]() не наложено никаких ограничений, позволило авторам дать следующую интерпретацию результатов: при эволюции сообщества к устойчивому нетривиальному равновесию (
не наложено никаких ограничений, позволило авторам дать следующую интерпретацию результатов: при эволюции сообщества к устойчивому нетривиальному равновесию (![]() ) скорость увеличения его равновесного разнообразия
) скорость увеличения его равновесного разнообразия ![]() должна быть по крайней мере не меньше скорости прироста общей численности (или биомассы).
должна быть по крайней мере не меньше скорости прироста общей численности (или биомассы).
4.13. Принцип максимального суммарного дыхания. Ф.Вашида (Washida, 1995) предложил гипотезу, согласно которой система стремится организовать свою биологическую конфигурацию так, чтобы максимизировать суммарное дыхание.
Для экосистемы, находящейся в стадии развития, доминирующим является рост как накопление суммарной биомассы, поэтому отношение P/R, где P – суммарная продукция экосистемы, а R – суммарное дыхание, значительно больше 1. При наступлении фазы зрелости, отношение P/R достигает 1. На этой стадии, которая является устойчивым состоянием, продукция достигает своего максимального уровня при данных условиях окружающей среды, а суммарное дыхание достигает максимального значения для данной продукции. Гипотеза максимального дыхания утверждает, что развитие экосистемы происходит так, чтобы как можно раньше достичь конфигурации, при которой дыхание максимально.
Была рассмотрена система, состоящая из двух биологических компонентов (продуцента и редуцента), единственного вида поступающей энергии (солнечная радиация), и одного питательного вещества. Продуцент потребляет солнечную энергию и ресурс, при этом увеличивает собственную биомассу и теряет тепло (дышит). Редуцент поглощает продуцента, при этом увеличивает свою биомассу, производит питательное вещество и дышит. Продуцирование и редуцирование синхронизированы во времени.
В модели приняты обозначения: x1 – количество биомассы, произведенной за некоторый период времени; x2 – количество питательного вещества, созданного за тот же период; e1, a21 – количество солнечной энергии и питательного вещества, необходимые для производства единицы продукции, соответственно; r1 – количество дыхания, генерируемое единицей продукции; a12 – количество биомассы, необходимое для производства единицы питательного вещества; b32 и r2 – соответственно масса редуцента и количество дыхания, созданное им при производстве единицы питательного вещества; p1, p3 – энергия, заключенная в единице биомассы продуцента и редуцента, соответственно; E – верхний предел солнечной радиации, которая может быть использована продуцентом; S – солнечная энергия, поступающая в систему, q – эффективность удержания этой энергии продуцентом.
Согласно первому закону термодинамики, должны выполняться следующие равенства: e1 = p1 + r1 и p1a12 = p3b32 + r2. Задача ставится следующим образом: максимизировать сумму R = r1x1 + r2x2 при условиях
e1x1 £ E = qS, a12x2 £ x1, a21x1 £ x2, 0 £ b32x2 и x1, x2 ³ 0.
Неравенства определяют область допустимых решений, среди которых ищется оптимальный набор (x1, x2). Условие, при котором существует хотя бы одно допустимое решение, имеет вид a12 a21 £ 1 – количество биомассы, косвенно необходимое для производства его же, должно быть меньше 1. Если это условие не выполнено, экосистема не может поддерживать состояние равновесия.
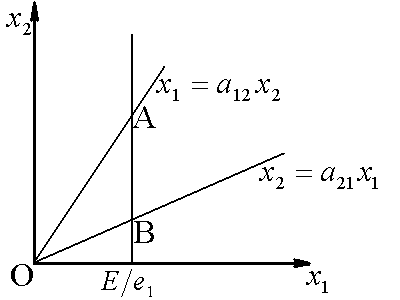 Область допустимых решений представлена на рис. 18 (треугольник АОВ).
Область допустимых решений представлена на рис. 18 (треугольник АОВ).
Рис.18. Область допустимых решений (OAB) задачи для системы продуцент – редуцент. x1 – количество биомассы, произведенной за некоторый период времени; x2 – количество питательного вещества, созданного за тот же период; e1, a21 – количество солнечной энергии и питательного вещества, необходимые для производства единицы продукции, соответственно; a12 – количество биомассы, необходимое для производства единицы питательного вещества; E – верхний предел солнечной радиации, которая может быть использована продуцентом
Поскольку величины r1 и r2 положительны, то требованию максимальности общего дыхания R удовлетворяет набор (x1, x2), соответствующий точке А. Биологически это означает, что редуцент перерабатывает всю биомассу, и питательное вещество возмещается в излишке.
Также была рассмотрена более сложная система, в которую входит два редуцента, потребляющих и выделяющих одни и те же вещества, только в разных соотношениях, т.е. их экологические ниши не идентичны, но сильно пересекаются. Питательные вещества, производимые редуцентами, полностью взаимозаменяемы.
Коэффициенты, относящиеся ко второму редуценту, обозначены буквами с верхним индексом “*”.
Энергетические соотношения в новой модели записываются равенствами e1 = p1+r1, p1a12 = p3b32+r2, p1a12* = p3b32*+r2*.
Задача максимального дыхания описывается следующими выражениями r1x1+r2x2+r2*x2*® max, e1x1 £ E, a12x2+a12*x2* £ x1, a21x1 £ x2+x2*, 0 £ b32x2+b32*x2*, x1, x2, x2* ³ 0.
Задача требует рассмотрения двух случаев.
1) Каждый из редуцентов способен в одиночку поддерживать систему. Математически это означает выполнение двух неравенств:
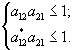
В этом случае решение задачи зависит от того, какой из редуцентов дает больший вклад в общее дыхание системы. Если ![]() >
> ![]() , то решение: x2 = 0,
, то решение: x2 = 0, ![]() > 0, иначе –x2 > 0,
> 0, иначе –x2 > 0, ![]() = 0 (равенство отношений означает эквивалентность разлагателей). После исключения одного из редуцентов задача сводится к предыдущей.
= 0 (равенство отношений означает эквивалентность разлагателей). После исключения одного из редуцентов задача сводится к предыдущей.
2) Новый редуцент не в состоянии в одиночку поддерживать систему, т е.
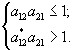
Если при этом он менее эффективен в выработке дыхания, то происходит его исключение из экосистемы. Содержательная ситуация имеет место при выполнении условия ![]() >
> ![]() .
.
 Область допустимых решений представляет собой пирамиду АВСО (рис.19).
Область допустимых решений представляет собой пирамиду АВСО (рис.19).
Рис.19. Область допустимых решений (OABC) задачи для системы продуцент – два редуцента. x1 – количество биомассы, произведенной за некоторый период времени; x2 – количество питательного вещества, созданного за тот же период; e1, a21 – количество солнечной энергии и питательного вещества, необходимые для производства единицы продукции, соответственно; a12 – количество биомассы, необходимое для производства единицы питательного вещества. Коэффициенты, относящиеся ко второму редуценту, обозначены буквами с верхним индексом “*”
Максимум дыхания достигается при наборе (x1, x2, x2*), соответствующем точке А. При таких значениях биомасс x1, x2, x2* солнечная энергия полностью перерабатывается и уровень производства питательного вещества и биомассы продуцента максимальны. Почему решением является именно точка А, ведь в этом случае первый редуцент должен снизить свой уровень активности “в угоду” новому и “в ущерб” себе? Рассматриваемая модель находится в устойчивом состоянии, согласно гипотезе максимального дыхания, система реорганизует свою конфигурацию для максимизирования дыхания, а это значит, что она стремится увеличить эффективность переработки энергии (на зрелом уровне это уже не накопление биомассы). Низкая эффективность использования энергии идентична уязвимости системы. Естественный отбор экосистем генерирует таких индивидов, которые объединяются для существования экосистемы даже ценой уменьшения численности их собственной популяции. Таким образом, стабильность экосистемы с максимальным уровнем дыхания гарантируется эволюцией.
На основании принципа максимального суммарного дыхания автор моделирует луговую экосистему. Модель состоит из 19 компонент и 16 видов энергетических и материальных потоков, связанных с компонентами. Компоненты разделены на 3 группы: продуценты, гетеротрофы — 3 вида травоядных и 3 вида плотоядных — и редуценты (12 составляющих). Энергетические и материальные потоки включают солнечную энергию, мертвую материю и другие аналогичные составляющие.
В результате исследования соответствующей вариационной задачи были получены следующие выводы:
Автор приходит к выводу, что используемая им модель достаточно адекватно описывает зрелые луговые экосистемы.
4.14. Принцип минимума общего осмотического давления. С.Шустер и Р.Гейнрих (Schuster, Heinrich, 1991) применяли метод условной экстремизации при изучении стационарного течения биохимических реакций. Они принимают, что общее осмотическое давление промежуточных продуктов в реакции можно записать в виде
![]() ,
,
где ![]() и
и ![]() обозначают, соответственно, концентрацию и коэффициент осмотического давления i-го промежуточного продукта реакции (зависимость
обозначают, соответственно, концентрацию и коэффициент осмотического давления i-го промежуточного продукта реакции (зависимость ![]() только от
только от ![]() является упрощающим предположением). Естественно, что осмотическое давление в клетке не может превосходить некоторой критической величины. Поэтому авторы считают, что при стационарном течении реакции, при котором все
является упрощающим предположением). Естественно, что осмотическое давление в клетке не может превосходить некоторой критической величины. Поэтому авторы считают, что при стационарном течении реакции, при котором все ![]() не зависят от времени,
не зависят от времени, ![]() должно принимать минимальное значение при ограничениях
должно принимать минимальное значение при ограничениях
![]() .
.
Здесь ![]() – некоторые равновесные константы реакции,
– некоторые равновесные константы реакции, ![]() – элементы стехиометрической матрицы реакционной системы. Ограничения – неравенства имеют смысл положительности скорости всех промежуточных реакций. Таким образом, при стационарном течении реакции, концентрации промежуточных продуктов
– элементы стехиометрической матрицы реакционной системы. Ограничения – неравенства имеют смысл положительности скорости всех промежуточных реакций. Таким образом, при стационарном течении реакции, концентрации промежуточных продуктов ![]() , находятся как решения следующей задачи
, находятся как решения следующей задачи
 (4.7)
(4.7)
В качестве примера С.Шустер и Р.Гейнрих рассматривают реакции метаболизма эритроцитов человека (Shuster, Heinrich, 1991). Поскольку эти реакции довольно сложные, то авторы получают лишь общую качественную картину решения задачи (4.7) в этом случае. Они отмечают, что эта картина согласуется с известными экспериментальными данными о метаболизме эритроцитов.
4.15. Принцип максимальной общей скорости биохимической реакции. Еще одно приложение метода условной оптимизации к исследованию биохимических реакций демонстрируют Т.Вильгельм с соавторами (Wilhelm et al., 1994). Они рассматривают биохимическую реакцию, состоящую из n элементарных шагов. Состояние реакции авторы характеризуют вектором ![]() , где
, где ![]() – константа скорости прямой (обратной) реакции i-го шага,
– константа скорости прямой (обратной) реакции i-го шага, ![]() . Авторы вводят гипотезу, что при стационарном течении общая скорость реакции
. Авторы вводят гипотезу, что при стационарном течении общая скорость реакции ![]() должна быть максимальна при ограничениях на величину
должна быть максимальна при ограничениях на величину ![]() и при фиксированной константе равновесия реакции
и при фиксированной константе равновесия реакции ![]() . Таким образом, для описания стационарного течения реакции авторы используют экстремальную задачу
. Таким образом, для описания стационарного течения реакции авторы используют экстремальную задачу
 (4.8)
(4.8)
Здесь ![]() и
и ![]() предполагаются известными параметрами задачи.
предполагаются известными параметрами задачи.
Авторы рассматривают несколько конкретных примеров, для которых результаты решения (4.8) согласуются с экспериментальными данными.
4.16. Принцип минимизации поверхностной энергии в развитии эмбриона. Н.Гоэл с соавторами (Goel et al., 1986) при помощи задачи на условный экстремум изучают один из ранних этапов развития эмбриона млекопитающих – процесс перехода между ранней и поздней морулой. Этот переход достаточно хорошо изучен. Он включает 4 этапа: клеточное деление, компактификацию, переферическое склеивание и появление внутренних клеток (интернализация). Компактификация состоит в том, что контактирующие части поверхностей клеток, возникших после деления и имеющих форму сфер, увеличиваются до такого состояния, что все внутриклеточное пространство, необходимо существующее между близко упакованными сферами, удаляется (путем индивидуальных деформаций первоначально сферических клеток), в то время как целый эмбрион приобретает сферическую форму. При этом компактификация идет именно до такого состояния, когда эмбрион приобретает форму сферы и не далее.
Для объяснения компактификации и интернализации авторы предлагают гипотезу, что клеточная конфигурация строится таким образом, что она минимизирует поверхностную энергию ![]() (
(![]() – число клеток в конфигурации) при условии фиксированного общего объема. Таким образом, получается задача на условный экстремум
– число клеток в конфигурации) при условии фиксированного общего объема. Таким образом, получается задача на условный экстремум
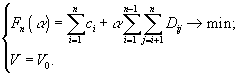 (4.9)
(4.9)
В выражении для поверхностной энергии ![]() в (4.9)
в (4.9) ![]() – площадь поверхности стенок i-ой клетки, находящихся в контакте только с окружающей средой (и не имеющих точек касания с другими клетками) и
– площадь поверхности стенок i-ой клетки, находящихся в контакте только с окружающей средой (и не имеющих точек касания с другими клетками) и ![]() – общая площадь стенок i-ой и j-ой клеток, V – объем. Параметр
– общая площадь стенок i-ой и j-ой клеток, V – объем. Параметр ![]() задается как некий весовой фактор для межклеточных поверхностей, отличающий их от стенок клеток, имеющих контакты только с внешней средой (для таких стенок параметр
задается как некий весовой фактор для межклеточных поверхностей, отличающий их от стенок клеток, имеющих контакты только с внешней средой (для таких стенок параметр ![]() считается равным единице).
считается равным единице).
Задача (4.9) сложна для аналитического решения. Поэтому авторы пытались решить ее численно для случая плоских (двухмерных) клеток и для некоторых специальных случаев трехмерных клеток (Lewis et al., 1988). Оказывается, что при ![]() минимум в (4.9) реализует конфигурация, при которой все клетки имеют форму кругов (или сфер) и каждая касается соседей только в одной точке, а при
минимум в (4.9) реализует конфигурация, при которой все клетки имеют форму кругов (или сфер) и каждая касается соседей только в одной точке, а при ![]() минимум в (4.9) реализует конфигурация, при которой каждая клетка деформируется таким образом, что внутриклеточные пространства исчезают, а вся совокупность клеток приобретает форму сферы (круга в двухмерном случае). Таким образом, по мнению авторов, процесс компактификации можно описать путем уменьшения
минимум в (4.9) реализует конфигурация, при которой каждая клетка деформируется таким образом, что внутриклеточные пространства исчезают, а вся совокупность клеток приобретает форму сферы (круга в двухмерном случае). Таким образом, по мнению авторов, процесс компактификации можно описать путем уменьшения ![]() в (1.5) с 2 до 0. (Физический механизм этого уменьшения авторы трактуют, связывая
в (1.5) с 2 до 0. (Физический механизм этого уменьшения авторы трактуют, связывая ![]() с концентрацией
с концентрацией ![]() ионов в окружающей эмбрион среде. Из экспериментальных наблюдений известно, что при компактификации эта концентрация уменьшается.)
ионов в окружающей эмбрион среде. Из экспериментальных наблюдений известно, что при компактификации эта концентрация уменьшается.)
Другим важным результатом численного исследования (4.9) оказывается тот факт, что при достаточно большом числе клеток ![]() конфигурация с внутренними клетками имеет меньшую энергию
конфигурация с внутренними клетками имеет меньшую энергию ![]() чем конфигурация без них (независимо от
чем конфигурация без них (независимо от ![]() ). По мнению авторов, это может служить возможным объяснением интернализации.
). По мнению авторов, это может служить возможным объяснением интернализации.
Таким образом, по мнению авторов, модель (4.9) достаточно адекватно описывает некоторые сложные процессы происходящие при развитии эмбриона млекопитающих.
4.17. Максимизация функции энтропии. В предыдущих пунктах обзора (см. п. 4.13 – 4.16) исследование биологической системы проводилось путем решения задачи на условный экстремум
 (4.10)
(4.10)
(здесь ![]() – некоторые фазовые переменные, описывающие состояние исследуемого объекта,
– некоторые фазовые переменные, описывающие состояние исследуемого объекта, ![]() – некоторая целевая функция (или функционал в более общем случае),
– некоторая целевая функция (или функционал в более общем случае), ![]() – функция состояния и
– функция состояния и ![]() – известные характеристики объекта,
– известные характеристики объекта, ![]() ), что позволяет сказать достаточно много об устройстве таких систем, исследование которых традиционным путем, например через решение некоторой системы дифференциальных уравнений, чрезвычайно трудоемко. Наименее разработанным этапом в данном подходе является нахождение целевой функции
), что позволяет сказать достаточно много об устройстве таких систем, исследование которых традиционным путем, например через решение некоторой системы дифференциальных уравнений, чрезвычайно трудоемко. Наименее разработанным этапом в данном подходе является нахождение целевой функции ![]() . (Ограничения
. (Ограничения ![]() обычно имеют смысл либо законов сохранения, либо прямо следуют из природы задачи.) Не случайно трудности нахождения или, говоря более точно, угадывания целевой функции особо отмечаются авторами всех вышеупомянутых работ. В некоторых работах приведены также ссылки на работы других авторов, которые при исследовании той же самой задачи используют другие целевые функции.
обычно имеют смысл либо законов сохранения, либо прямо следуют из природы задачи.) Не случайно трудности нахождения или, говоря более точно, угадывания целевой функции особо отмечаются авторами всех вышеупомянутых работ. В некоторых работах приведены также ссылки на работы других авторов, которые при исследовании той же самой задачи используют другие целевые функции.
В 1957 году Э.Джейнс, основываясь на внешнем сходстве формулы Гиббса для энтропии и формулы Шеннона для средней информации на символ в сообщении, предложил “Формализм максимальной энтропии” (The Maximum Entropy Formalism) для исследования сложных систем (Jaynes, 1957а,b).
Вкратце последовательность применения формализма Джейнса следующая (Трайбус, 1970):
1) перечислить все микросостояния системы и обозначить символом ![]() вероятности каждого из них,
вероятности каждого из них, ![]() ;
;
2) перечислить все, что известно о “средних значениях” величин, связанных с системой и выразить средние значения в виде следующих уравнений:
![]() ,
,
где xi – значение признака X,свойственное состоянию i; gr(X) – функция от X; ![]() – известное среднее значение;
– известное среднее значение;
3) максимизировать энтропию
![]()
используя уравнения для средних значений и дополнительное условие нормировки
![]()
и таким образом найти ![]() , которые “реализуются в действительности”.
, которые “реализуются в действительности”.
Вероятность ![]() понимается в широком смысле, то есть
понимается в широком смысле, то есть ![]() может быть, например, относительным числом элементов системы, обладающих некоторым признаком
может быть, например, относительным числом элементов системы, обладающих некоторым признаком ![]() .
.
Таким образом, Э.Джейнс предлагает в качестве целевой функции использовать энтропию ![]() .
.
Э.Джейнс и его последователи показали, что с помощью формализма максимальной энтропии можно легко получить все основные соотношения классической термодинамики и исследовать другие физические системы (The maximum entropy formalism, 1978).
Г.Хакен (Хакен, 1991) успешно применил формализм Джейнса в теории неравновесных фазовых переходов, в том числе и к некоторым биологическим системам, в которых наблюдаются аналоги таких переходов.
Эффективное применение вариационных принципов в статистической физике и в теории информации в виде условной максимизации функции энтропии для характеристики стационарного состояния сложной системы дало толчок к использованию подобного инструмента для решения некоторых биологических проблем. В частности, А.К.Приц рассматривает популяцию как открытую термодинамическую систему, стремящуюся сохранить свою структуру, описываемую функцией энтропии, неизменной во времени (Приц, 1974). В работах Д.Лурье с соавторами (Lurie et al., 1983) предлагается условная максимизация энтропиеподобной функции для описания адаптивного поведения сложной системы. (Следует отметить, что авторы указанных работ не ссылаются на Э.Джейнса и, по-видимому, не знакомы с его работами. Они используют идеи статистической физики (подход, на плодотворность которого в биологии указывал, в частности, Ю.М.Свирежев (1991)), но, как будет видно из дальнейшего, их метод полностью укладывается в рамки формализма Джейнса.)
А.К.Приц (1974), изучая возрастную структуру популяции рыб, видел большое значение подобных исследований для рыболовного промысла (предсказание будущих уловов и предотвращение переуловов).
В качестве статистической характеристики состояния популяции, по которой можно судить об изменениях возрастной структуры, А.К.Приц предложил энтропию S: ![]() , где
, где ![]() Эти обозначения соответствуют численности популяции (N), числу особей возраста i лет (Ni), предельному возрасту особей популяции (n). Вклад каждой возрастной группы в общую численность представляет собой вероятность pi встретить особь возраста i лет среди N особей популяции.
Эти обозначения соответствуют численности популяции (N), числу особей возраста i лет (Ni), предельному возрасту особей популяции (n). Вклад каждой возрастной группы в общую численность представляет собой вероятность pi встретить особь возраста i лет среди N особей популяции.
Популяция рассматривается как “открытая термодинамическая система, находящаяся в состоянии непрерывного обмена с окружающей средой, самовоспроизводящаяся и саморегулирующаяся”. А.К.Приц предлагает исходить из принципа стационарного состояния открытых систем, согласно которому все живые системы стремятся сохранить свою структуру (и энтропию) неизменной во времени.
Энтропия популяции записывается в виде ![]() общая фиксированная численность задается выражением
общая фиксированная численность задается выражением ![]() , а фиксированная масса популяции – выражением
, а фиксированная масса популяции – выражением ![]() (mi – масса одной особи i-го возраста).
(mi – масса одной особи i-го возраста).
Задача нахождения условий, при которых осуществляется стационарное состояние, удовлетворяющее перечисленным выше условиям, решается применением метода неопределенных множителей Лагранжа к системе
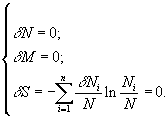
В результате была получена формула, определяющая связь между возрастной численностью i-й группы Ni, массой mi отдельной особи i-го возраста и общей численностью популяции N
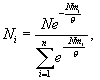 (4.11)
(4.11)
где θ – модуль статистического распределения различных особей по возрастам, который определяется из эксперимента на основе физических соображений.
Формула (4.11) может быть представлена в несколько ином виде, если в качестве фиксированного параметра взять среднее время жизни популяции Т, ![]() (ti – возраст i-й группы популяции)
(ti – возраст i-й группы популяции)
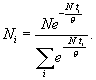 (4.12)
(4.12)
Для практического применения формулы (4.12) интересным представляется вид параметра θ. А.К.Приц получил соотношение θ = Nm1, справедливое для долгоживущих рыб. Такой вид параметра θ обеспечивает оптимальности пополнения популяции, т.е. численности N1 (в условиях интенсивного вылова воспроизводство должно быть максимальным). При выводе указанного соотношения предполагалось, что система находится в стационарном состоянии. Формула (4.11) может быть теперь записана как 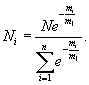 Зная по результатам экспериментального лова массу mi особи i-го возраста, а также число особей Ni, можно найти общую численность популяции N и остальные численности Nj, общую массу популяции. Были проведены расчеты для сельди Северного моря с 1947 по 1971 год. Сравнение расчетных и реальных значений дало совпадение от 70% и выше за каждый год, кроме одного.
Зная по результатам экспериментального лова массу mi особи i-го возраста, а также число особей Ni, можно найти общую численность популяции N и остальные численности Nj, общую массу популяции. Были проведены расчеты для сельди Северного моря с 1947 по 1971 год. Сравнение расчетных и реальных значений дало совпадение от 70% и выше за каждый год, кроме одного.
Предсказание численностей различных возрастных групп дает возможность рассчитывать ежегодный прирост биомассы по всей популяции
![]()
что, в свою очередь, позволяет регулировать промысел для поддержания стабильного значения биомассы. Причем найденный прирост ∆М и определяет максимально возможный вылов в соответствии со стратегией оптимального вылова.
По мнению Д.Лурье и его коллег (Lurie et al., 1983) живым системам присуще адаптивное поведение, которое обусловлено конкуренцией между двумя тенденциями – тенденцией к большему разнообразию, происходящей из постоянного появления ошибок в генетическом коде, и тенденцией к меньшему разнообразию из-за естественного отбора. Для математического выражения этой конкуренции предлагается ввести понятие удельного разнообразия биомассы ![]() , которое находится в тесной связи с физическими параметрами экосистемы
, которое находится в тесной связи с физическими параметрами экосистемы
![]()
где p(m) – плотность вероятности распределения биомассы. В качестве задачи ставится нахождение функции плотности распределения биомассы, максимизирующей разнообразие ω при ограничении на полную биомассу M, которую может поддерживать эта система ![]() где N – количество особей данной системы,
где N – количество особей данной системы, ![]() – средняя биомасса особи.
– средняя биомасса особи.
Итак, рассматривается вариационная задача
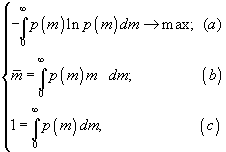
здесь в (а) для удобства используется натуральный логарифм, (b) представляет собой запись ограничения в терминах средней биомассы, (с) – условие нормировки распределения p(m).
Решение задачи методом неопределенных множителей Лагранжа дает следующую экспоненциально убывающую функцию биомассы
![]()
Также было предложено дискретное рассмотрение проблемы. Область значений биомассы разделяется на r интервалов размером ![]() . В этом случае разнообразие биомассы определяется выражением
. В этом случае разнообразие биомассы определяется выражением
![]()
где ni – число особей экосистемы в i-м интервале, независимо от вида, ![]() (для больших популяций
(для больших популяций ![]() ).
).
Введя обозначение ![]() и записав в дискретной форме ограничение и условие нормировки, получим систему
и записав в дискретной форме ограничение и условие нормировки, получим систему
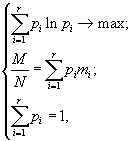
где mi – представительное значение биомассы особей из i-го интервала.
Решение данной вариационной задачи задается формулой: 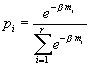 . Для нахождения множителя Лагранжа β и знаменателя дроби в выражении для относительной численности pi необходим переход к пределу и интегрирование (
. Для нахождения множителя Лагранжа β и знаменателя дроби в выражении для относительной численности pi необходим переход к пределу и интегрирование (![]() ,
,![]() ). Проделав необходимые операции, получим уже выведенную ранее формулу
). Проделав необходимые операции, получим уже выведенную ранее формулу ![]()
Описанные теоретические результаты были проверены на различных данных о распределениях рыб по весам в уловах. Коэффициент корреляции изменялся от 0,986 до 0,998. Рассмотрение дискретной записи задачи предполагает зависимость разнообразия ω от разбиения биомассы экосистемы на классы ![]() . Для данных одной из экспедиций были проведены расчеты с различными
. Для данных одной из экспедиций были проведены расчеты с различными ![]() , при этом были получены значения коэффициентов корреляции 0,971; 0,979; 0,986.
, при этом были получены значения коэффициентов корреляции 0,971; 0,979; 0,986.
Следует отметить недостаток предложенной теории, на который обратил внимание Дж.Розен (Rosen, 1984) в своей работе “Замечание по поводу разнообразия биомассы в экологии”: “…в постулате Лурье-Вагенсберга все ячейки биомассы размера ![]() a priori обладают одинаковым статистическим весом, хотя такое предположение об эквивалентности
a priori обладают одинаковым статистическим весом, хотя такое предположение об эквивалентности ![]() -ячеек противоречит соображениям эргодичности, основанным на динамике популяции”.
-ячеек противоречит соображениям эргодичности, основанным на динамике популяции”.
В качестве альтернативы Дж.Розеном был предложен динамический подход. Рассматривается уравнение динамической непрерывности для распределения биомассы в большой популяции рыб:
![]()
Здесь m – биомасса индивидуума, n = n(t,m) – число индивидуумов на единичный интервал биомассы, ![]() – положительная средняя скорость накопления биомассы,
– положительная средняя скорость накопления биомассы, ![]() – положительная средняя дисперсионная скорость для n, обусловленная генетическими различиями и различиями в условиях окружающей среды, а
– положительная средняя дисперсионная скорость для n, обусловленная генетическими различиями и различиями в условиях окружающей среды, а ![]() – положительная средняя удельная скорость, с которой индивидуумы удаляются из популяции. Величины
– положительная средняя удельная скорость, с которой индивидуумы удаляются из популяции. Величины ![]() аппроксимируются усредненными по времени и массе постоянными. Общее решение динамического уравнения с учетом указанных ограничений приводит к асимптотическому стационарному уравнению:
аппроксимируются усредненными по времени и массе постоянными. Общее решение динамического уравнения с учетом указанных ограничений приводит к асимптотическому стационарному уравнению: ![]() , где численность n удовлетворяет уравнению
, где численность n удовлетворяет уравнению
 (4.13)
(4.13)
Решение уравнения (4.13) задается формулой ![]() где А – свободная константа, зависящая от начального значения численности n = n(t,m), а значение
где А – свободная константа, зависящая от начального значения численности n = n(t,m), а значение ![]() определяется уравнением
определяется уравнением  .
.
Вернемся к теории Лурье-Вагенсберга. Как уже отмечалось выше, результаты, полученные по выведенным формулам, хорошо согласуются с некоторыми опытными данными. Однако, это касается только тех популяций, в которых нет прямого взаимодействия между различными классами биомассы, что соответствует использованию в теоретическом выводе независимости вероятностей pi. В частности, гипотеза оказалась полностью несостоятельной при описании популяций хека, который характеризуется каннибалистическим поведением. В работе Дж.Вагенсберга и Дж.Валлса (Wagensberg, Valls, 1987) была предложена модификация принципа максимума энтропии, описывающая внутреннее взаимодействие популяции “хищник-жертва”.
Популяция предполагается состоящей из двух взаимодействующих классов, каждый из которых внутренне независим. Пусть ![]() – вероятность нахождения хищника в j-м классе биомассы, p(mi) – вероятность нахождения жертвы в i-м классе биомассы,
– вероятность нахождения хищника в j-м классе биомассы, p(mi) – вероятность нахождения жертвы в i-м классе биомассы, 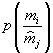 – условная вероятность, связанная соотношением
– условная вероятность, связанная соотношением 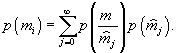 Энтропия теперь выглядит следующим образом
Энтропия теперь выглядит следующим образом
![]() , где
, где  . (4.14)
. (4.14)
Предлагаются следующие гипотезы:
1) внутренне взаимодействие определенной популяции характеризуется определенной матрицей взаимодействия, для элементов которой выполнено
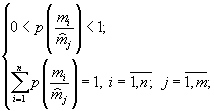
2) устойчивая статистическая структура для данной матрицы взаимодействия ![]() максимизирует объединенную энтропию хищника и жертвы при ограничениях
максимизирует объединенную энтропию хищника и жертвы при ограничениях
 (4.15)
(4.15)
При решении методом неопределенных множителей Лагранжа вариационной задачи, составленной из уравнений (4.14, 4.15) (![]() ) было получено распределение биомассы
) было получено распределение биомассы
![]() . (4.16)
. (4.16)
Значение Z и β – множитель Лагранжа – вычисляются при подстановке (4.16) в (4.15). Структура “жертвы” вычисляется с помощью формулы для величины p(m).
Для проверки модифицированной гипотезы была рассмотрена популяция хека, состоящая из особей возраста от одного года до 13 лет. Соответственно было выделено 13 классов массы. Особи первых 7-ми классов предполагались жертвами, остальные, с 8-го по 13-й – хищниками, т.е. уравнение для p(m) имело вид
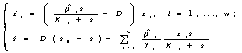
(все массы ![]() и матрица
и матрица ![]() – известны).
– известны).
Сравнение вычисленных и наблюдаемых распределений показало, что различие в коэффициенте β составляет 0,71% против 70% для гипотезы, не учитывающей эффект каннибализма, а коэффициент корреляции составил 0,999.
Применение современных методов вариационного моделирования в экологии сообществ содержится в работах А.П.Левича с коллегами (Левич, 1980; Левич и др., 1997; Levich, 2000). Моделируется сообщество одноклеточных организмов, потребляющих ресурсы, которые не могут заменить друг друга, поскольку выполняют различные функции по отношению к росту. Допустимо деление клеток, смертность, но не их слияние. В лабораторных условиях описываемая модель соответствует накопительному культивированию, при котором не происходит добавление или изъятие ресурсов и микроорганизмов. Изучается развитие поликультуры до остановки, вызванной исчерпанием одного из ресурсов, но не какими-либо иными причинами.
Постулируется, что динамические системы из заданного состояния переходят в состояние с экстремальной (в пределах, допустимых имеющимися ресурсами) структурой. Соответствующая вариационная задача на условный экстремум выглядит следующим образом (Левич и др., 1997; Levich, 2000):
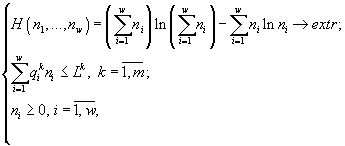 (4.17)
(4.17)
где ni – конечная искомая численность каждого из видов, ![]() – количество k-ого ресурса, необходимое для роста вида i, в расчете на одну клетку, называемое потребностью вида i в ресурсе k, m – общее количество взаимонезаменимых ресурсов, потребляемых сообществом, w – число видов в сообществе, Lk – начальное содержание ресурса k в среде (Lk
– количество k-ого ресурса, необходимое для роста вида i, в расчете на одну клетку, называемое потребностью вида i в ресурсе k, m – общее количество взаимонезаменимых ресурсов, потребляемых сообществом, w – число видов в сообществе, Lk – начальное содержание ресурса k в среде (Lk ![]() ) Отметим, что условия, ограничивающие экстремум, имеют форму неравенств, а не равенств.
) Отметим, что условия, ограничивающие экстремум, имеют форму неравенств, а не равенств.
![]() Важно также отметить, что функционал
Важно также отметить, что функционал ![]() , названный обобщенной энтропией, не постулируется, а выводится (Левич, 1982) на основе категорно-функторного метода сравнения математических структур. Можно следующим образом описать нахождение целевой функции, максимум которой дает искомое стационарное состояние (Левич, 1982; Levich, 1995; Levich, Solov'yov, 1999).
, названный обобщенной энтропией, не постулируется, а выводится (Левич, 1982) на основе категорно-функторного метода сравнения математических структур. Можно следующим образом описать нахождение целевой функции, максимум которой дает искомое стационарное состояние (Левич, 1982; Levich, 1995; Levich, Solov'yov, 1999).
Для формального описания исследуемой нами системы следует выбрать определенную математическую структуру. Далее необходимо ввести математическую категорию ![]() , объекты которой соответствуют всевозможным состояниям нашей системы и являются множествами с выбранной структурой, а морфизмы соответствуют допускаемым природой системы переходам из одного состояния в другое, и являются сохраняющими математическую структуру преобразованиями структурированных множеств.
, объекты которой соответствуют всевозможным состояниям нашей системы и являются множествами с выбранной структурой, а морфизмы соответствуют допускаемым природой системы переходам из одного состояния в другое, и являются сохраняющими математическую структуру преобразованиями структурированных множеств.
Параллельно вводится категория ![]() . Объекты в категории
. Объекты в категории ![]() те же, что и в категории Q, но со "стертой" структурой, то есть рассматриваемые просто как множества. Морфизмы в категории
те же, что и в категории Q, но со "стертой" структурой, то есть рассматриваемые просто как множества. Морфизмы в категории ![]() обладают теми же свойствами, что и в категории Q, но без требования сохранения структуры.
обладают теми же свойствами, что и в категории Q, но без требования сохранения структуры.
Поскольку каждый допустимый структурой системы морфизм переводит систему в новое состояние без изменения общей структуры, то число сохраняющих структуру морфизмов можно интерпретировать как число "микросостояний" системы, т.е. общую структуру. Такая интерпретация подсказывает, что логарифм удельного числа преобразований системы связан с больцмановским определением энтропии в статистической механике.
Поэтому для каждого состояния ![]() исследуемой системы, то есть для каждого объекта в категории Q вводится величина, называемая обобщенной энтропией состояния A,
исследуемой системы, то есть для каждого объекта в категории Q вводится величина, называемая обобщенной энтропией состояния A,
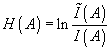 . (4.18)
. (4.18)
В формуле (4.18) величина![]() есть мощность множества
есть мощность множества ![]() морфизмов категории
морфизмов категории ![]() из A в само себя и аналогично величина
из A в само себя и аналогично величина![]() есть мощность множества
есть мощность множества ![]() . Отметим, что по определению понятия категории множество
. Отметим, что по определению понятия категории множество ![]() , так как содержит хотя бы единичный морфизм.
, так как содержит хотя бы единичный морфизм.
Возвращаясь к вариационной задаче (4.17), отметим, что здесь сообщество описывается математической структурой множеств из n элементов, разбитых на w непересекающихся классов c числом элементов ni (i = 1,…,w) в каждом.
Именно энтропию (4.18) предлагается рассматривать как целевую функцию в задаче (4.10).
Формулу (4.18) для энтропии можно интерпретировать как меру структурированности состояния A, то есть меру отклонения структуры состояния A от его бесструктурного аналога. Таким образом, согласно экстремальному принципу (4.10) осуществляется состояние системы, которое сильнее других удалено от своего полностью бесструктурного прообраза, т.е. максимально структурировано.
Обсуждаемый экстремальный принцип постулируется как реализация состояний системы с экстремальной структурой. Конструкция сравнения структурированных множеств (Левич, 1982; Levich, Solov’yov,1999; Левич, 2001) аналогична конструкции сравнения бесструктурных множеств по мощностям. Поэтому предложенный экстремальный принцип обобщает на многовидовые сообщества принцип максимальной экспансии (в пределах доступных ресурсов), подразумеваемый обычно в биологии для отдельных популяций.
Энтропийному принципу эквивалентны некоторые термодинамические экстремальные принципы (Levich, 2000). Максимум энтропии H с ограничением по энергии Е влечёт минимум функции Лагранжа F=E - TH, называемой в термодинамике свободной энергией. (Поясним связь абсолютной температуры T с множителем Лагранжа. Подстановка решения задачи (4.17), о котором подробнее будет сказано ниже, в формулу для энтропии представляет энтропию как функцию лимитирующих развитие системы ресурсов ![]() . В термодинамическом случае, когда речь идет о максимуме энтропии газа при ограничении только потоком тепла
. В термодинамическом случае, когда речь идет о максимуме энтропии газа при ограничении только потоком тепла ![]() , получаем
, получаем![]() , где
, где ![]() – соответствующий множитель Лагранжа. Тем самым устанавливается связь “обобщенной энтропии” H с энтропией феноменологической термодинамики.)
– соответствующий множитель Лагранжа. Тем самым устанавливается связь “обобщенной энтропии” H с энтропией феноменологической термодинамики.)
Описываемый экстремальный принцип непосредственно связан с принципом максимума эксергии, о котором уже говорилось в п. 4.9. Напомним понятие эксергии и сформулируем связанный с ним принцип максимума несколько другими словами. Величина Ex = T(Heq - H), где Heq – энтропия системы в состоянии термодинамического равновесия, называется в термодинамике эксергией (т.е. энергией, которая может быть преобразована в полезную работу при участии заданной окружающей среды). С.Йоргенсен (Jørgensen, 1986; 1988; Jørgensen et al., 1995) в контексте экологического моделирования выдвинул и результативно применил принцип максимума эксергии: изменения воздействий на систему будут вызывать изменения структуры системы, которые сопровождаются неотрицательными изменениями эксергии. Величина эксергии с точностью до множителя совпадает с экстремизируемой в данном подходе “обобщённой энтропией”. Таким образом, предложенный в данной работе экстремальный принцип представляет собой модификацию принципа максимальной эксергии С.Йоргенсена, дополненную нетермодинамическим методом расчёта энтропии (Левич, 1982).
Еще одна интерпретация энтропийного экстремального принципа связана с, так называемой, теоремой Гиббса. Можно показать (Левич, Алексеев, 1997; Левич, Фурсова, 2003), что задача
для любого ![]() равносильна задачам
равносильна задачам

Таким образом, оказывается, что вместо формулировки экстремального принципа “энтропия системы максимальна, при условии, что потребление ресурсов ниже определённого запаса” справедлива формулировка “потребление системой ресурсов минимально, при условии, что энтропия системы (или её сложность, разнообразие, и т.д.,) выше некоторого порога”, т.е. принцип максимума энтропии эквивалентен принципу минимума потребления системой лимитирующих ресурсов. Указанная теорема является обобщением теоремы Гиббса (Gibbs, 1902) о равносильности требования максимальности энтропии газа (при заданной энергии) процедуре минимизации энергии (при заданной энтропии).
Кроме того, показано (Левич, 1978), что энтропию (4.18) можно рассматривать как количество информации, связанной со структурой системы. Поэтому экстремальность энтропии можно интерпретировать и в информационных терминах.
Величину ![]() (
(![]() ) экологи применяют как индекс разнообразия видовой структуры сообществ (McArthur, 1955). Поэтому рассматриваемый экстремальный принцип может интерпретироваться как требование максимального видового разнообразия сообществ (см., например, Lurie et al., 1983).
) экологи применяют как индекс разнообразия видовой структуры сообществ (McArthur, 1955). Поэтому рассматриваемый экстремальный принцип может интерпретироваться как требование максимального видового разнообразия сообществ (см., например, Lurie et al., 1983).
Основным результатом, на котором базируется последующее исследование сформулированной задачи, является теорема стратификации. Приводим ее формулировку. Все пространство ресурсных факторов ![]() распадается (стратифицируется) на
распадается (стратифицируется) на ![]() непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте SJ, где J — непустое подмножество множества ресурсов {1,2,...,m}, выполняется:
непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте SJ, где J — непустое подмножество множества ресурсов {1,2,...,m}, выполняется:
1) решение задачи (4.17) ![]() , где
, где ![]() º
{L1, L2,..., Lm}, зависят только от тех Lk, для которых kÎ
J;
º
{L1, L2,..., Lm}, зависят только от тех Lk, для которых kÎ
J;
2) на этом решении нестрогие неравенства ![]() обращаются в строгие равенства для всех kÎ
J и в строгие неравенства для всех kÏ
J;
обращаются в строгие равенства для всех kÎ
J и в строгие неравенства для всех kÏ
J;
Теорема стратификации влечёт редукцию задачи (4.17) к задачам
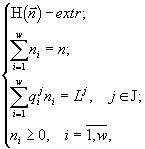 (4.19)
(4.19)
формулируемым для любого J Ì {1,2,...,m} (Левич и др., 1994).
Биологическая интерпретация теоремы стратификации задаёт правило лимитирующего звена для многовидовых сообществ, популяции из которых конкурируют за несколько ресурсов (Левич и др., 1993б). Лимитирующими называются ресурсы, потребляемые сообществом из среды полностью, т.е. те, для которых балансовые неравенства задачи (4.17) превращаются в равенства. Согласно теореме стратификации, из заданной совокупности ресурсов ![]() полностью потребляются, т.е. лимитируют рост сообщества, ресурсы из набора J, идентифицирующего страт SJ, которому принадлежит заданный вектор
полностью потребляются, т.е. лимитируют рост сообщества, ресурсы из набора J, идентифицирующего страт SJ, которому принадлежит заданный вектор ![]() . Таким образом, теорема стратификации позволяет строго предсказывать ресурсы, лимитирующие рост сообщества с заданным набором потребностей
. Таким образом, теорема стратификации позволяет строго предсказывать ресурсы, лимитирующие рост сообщества с заданным набором потребностей ![]() .
.
Для монокультур правило лимитирующего звена совпадает с принципом минимума Либиха (Liebig, 1840), а именно, при выполнении условия ![]() рост лимитирован фактором 1. Указанное условие эквивалентно неравенству
рост лимитирован фактором 1. Указанное условие эквивалентно неравенству ![]() , т.е. лимитирующим оказывается ресурс, для которого достигаемая на его запасе численность минимальна (т.е. тот, который в только что указанном смысле “находится в минимуме”). Формально задача (4.17) не определена для w = 1, поскольку функционал в этом случае обращается в константу. Но решение задачи в этом случае может быть получено только из ресурсных ограничений, согласно которым
, т.е. лимитирующим оказывается ресурс, для которого достигаемая на его запасе численность минимальна (т.е. тот, который в только что указанном смысле “находится в минимуме”). Формально задача (4.17) не определена для w = 1, поскольку функционал в этом случае обращается в константу. Но решение задачи в этом случае может быть получено только из ресурсных ограничений, согласно которым
![]() ,
,
т.е. реализуется численность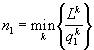 , или принцип минимума Либиха.
, или принцип минимума Либиха.
Для неодновидовых сообществ согласно правилу лимитирующего звена существуют как области, где рост ограничен одним фактором, так и несколькими, в частности, всеми факторами. Последняя ситуация в агрохимической литературе носит название “закона совокупного действия факторов” Митчерлиха (Mitscherlich, 1909). Эксперименты с фитопланктоном (Droop, 1973; Rhee, 1978; Ahlgren, 1980; Groot, 1983) демонстрируют предсказываемое переключение лимитирования. Результаты подробного изучения некоторых конкретных примеров одно-, двух и трехфакторного лимитирования представлены в книге А.П.Левича с соавторами (Левич и др., 1997).
Решение задач (4.19) получило название формулы видовой структуры (Левич, 1980)
![]()
где вектора ![]() ,
, ![]() и
и ![]() имеют компоненты j из набора J, идентифицирующего страт, которому принадлежит вектор
имеют компоненты j из набора J, идентифицирующего страт, которому принадлежит вектор ![]() . Множители Лагранжа
. Множители Лагранжа ![]() и полная численность n как функции потребляемых полностью в страте SJ ресурсов
и полная численность n как функции потребляемых полностью в страте SJ ресурсов ![]() ищутся из алгебраических уравнений
ищутся из алгебраических уравнений

Для проверки адекватности модели был проведен ряд экспериментов (Левич и др., 1997). В опыте с поликультурой из 10-ти видов зелёных микроводорослей (Chlorella vulgaris (Bejerink.), Scotiella nivalis (Fritsch.), Chromochloris cinnoborina (Chodat.), S. quadricauda (Turp.), Scenedesmus bijugatus (Lagerh.), Scenedesmus obliquus (Kruger.), Ankistrodesmus acicularis (Korschik.), A. braunii (Brunnth.), Stichococcus mirabilis (Lagerh.) и Chlamydomonas humicola (Luksch.)) в течение 70 суток в нескольких повторностях измеряли численности и биомассы видов, первичную продукцию, концентрации минеральных форм азота и фосфора. В монокультурах тех же видов измеряли потребности клеток в фосфоре. Содержание биогенных элементов в среде было подобрано так, чтобы развитие водорослей ограничивалось фосфором среды. На стационарной фазе роста распределение численности клеток описывалось формулой видовой структуры ![]() с коэффициентом множественной корреляции 0,97 (Левич и др. 1986). Эта же серия опытов наряду с ещё шестнадцатью сериями экспериментов была проанализирована с помощью ранговых критериев адекватности выборок и с помощью калибровки клеточных квот (Замолодчиков и др., 1993). Серии опытов отличались друг от друга наборами таксонов (от четырех до десяти видов в поликультуре) и начальными концентрациями азота и фосфора в среде (Левич и др., 1997). При лимитировании роста единственным ресурсом место вида в ранжированном по убыванию ряду численностей на стационарной фазе согласно формуле видовой структуры полностью определяется рангом клеточной квоты вида по лимитирующему ресурсу (наибольшая численность соответствует наименьшей клеточной потребности). Была сделана попытка подобрать в пределах экспериментальной ошибки единственный набор рангов измеренных в опыте потребностей так, чтобы воспроизвести экспериментальные ранги численностей уже во всех сериях, где рост был ограничен единственным ресурсом. Это удалось сделать с ранговым коэффициентом корреляции Спирмена в пределах от 0,92 до 1,00. Для серий с неоднофакторным лимитированием процедура проверки адекватности формулы видовой структуры состояла в следующем. По численностям видов в паре серий рассчитывали потребности по азоту и фосфору, а для всех остальных серий с этими откалиброванными потребностями по формуле видовой структуры рассчитывали численности видов и сравнивали их с экспериментальными значениями. Сравнение показало, что рассчитанные значения численностей совпадают с измеренными в опыте в пределах экспериментальных ошибок.
с коэффициентом множественной корреляции 0,97 (Левич и др. 1986). Эта же серия опытов наряду с ещё шестнадцатью сериями экспериментов была проанализирована с помощью ранговых критериев адекватности выборок и с помощью калибровки клеточных квот (Замолодчиков и др., 1993). Серии опытов отличались друг от друга наборами таксонов (от четырех до десяти видов в поликультуре) и начальными концентрациями азота и фосфора в среде (Левич и др., 1997). При лимитировании роста единственным ресурсом место вида в ранжированном по убыванию ряду численностей на стационарной фазе согласно формуле видовой структуры полностью определяется рангом клеточной квоты вида по лимитирующему ресурсу (наибольшая численность соответствует наименьшей клеточной потребности). Была сделана попытка подобрать в пределах экспериментальной ошибки единственный набор рангов измеренных в опыте потребностей так, чтобы воспроизвести экспериментальные ранги численностей уже во всех сериях, где рост был ограничен единственным ресурсом. Это удалось сделать с ранговым коэффициентом корреляции Спирмена в пределах от 0,92 до 1,00. Для серий с неоднофакторным лимитированием процедура проверки адекватности формулы видовой структуры состояла в следующем. По численностям видов в паре серий рассчитывали потребности по азоту и фосфору, а для всех остальных серий с этими откалиброванными потребностями по формуле видовой структуры рассчитывали численности видов и сравнивали их с экспериментальными значениями. Сравнение показало, что рассчитанные значения численностей совпадают с измеренными в опыте в пределах экспериментальных ошибок.
Важным свойством формулы видовой структуры является зависимость относительных численностей только от отношений ресурсов, что позволяет управлять видовыми обилиями с помощью отношений ресурсов в среде. Доказано (Левич и др., 1993а; Alexeyev, Levich, 1997), что относительная численность некоторого вида принимает наибольшее возможное значение при отношениях ресурсов в среде, равных отношениям потребностей в этих ресурсах у данного вида. Эта гипотеза была испытана в традиционной имитационной модели процессов потребления и роста для фитопланктона (Левич, Личман, 1992; см. также п.3.5). Законы Моно, Друпа и сохранения вещества, использованные в модели, являются эмпирическими обобщениями. Тем самым вся модель работает как "большая аппроксимирующая формула", аккумулирующая опытные знания. Машинные эксперименты с моделью показали, что в этих накопленных десятилетиями знаниях неявно заложены эффекты зависимости относительных обилий популяций сообщества от исходных отношений количеств ресурсов в среде. При этом оптимальные для каждого вида отношения оказались близки к отношениям минимальных клеточных квот для этого вида. Другими словами, формальная теорема вариационного моделирования была подтверждена современными представлениями о функционировании фитопланктонных сообществ, заложенными в традиционную феноменологическую модель.
Кроме того, гипотеза о роли отношений концентраций питательных веществ была проверена в ряде экспериментов по регуляции состава альгоценозов с лабораторными поликультурами и с сообществами естественного фитопланктона in vitro и in situ (Левич и др., 1992; 1996а,б; 1997; Левич, Булгаков, 1993; Levich, Bulgakov, 1992; Bulgakov, Levich, 1999).
Многочисленные серии лабораторных экспериментов (Левич, Булгаков, 1993; Левич и др., 1997) отличались друг от друга составом альгоценозов и начальными концентрациями биогенных веществ в среде. В повторностях измеряли численности, объёмы клеток; концентрации ресурсов в среде как в поликультурах, так и в соответствующих монокультурах. Контролировали начальные внутриклеточные запасы лимитирующих ресурсов; нелимитирующие параметры: свет, температуру, кислотность, микроэлементы; альгологическую и бактериологическую чистоту, отсутствие метаболического взаимодействия. Рассчитывали потребления всем сообществом ресурса и клеточные потребности. Опыты в режиме накопительного культивирования продолжались до стационарной стадии роста. Отметим, что ни один из проведённых опытов не противоречил следствиям вариационного моделирования.
С естественным прудовым фитопланктоном in vitro проводились два опыта в различные сезоны (Левич и др., 1992). Прудовое сообщество содержало более сотни видов, принадлежащих к нескольким крупным таксонам: Chlorophyta (Chlorococcales, Chlamydomonadales, Desmidiales), Bacillariophyta, Cyanophyta. В одной из серий (20 литровые аквариумы) были испытаны три различных отношения азота к фосфору в интервале от 5 до 16, в другой серии (двухлитровые колбы) – пять отношений в интервале от 2 до 200. Опыты проводились в нескольких повторностях. Основная тенденция состояла в следующем: для протококковых водорослей оптимальны высокие отношения N/P, а для синезелёных и диатомовых – низкие (на отношение N/P реагируют, конечно, не высшие таксоны, а доминирующие в них виды: в Chlorococales это S. quadricauda, в Cyanophyta – виды рода Microcystis, в Bacillariophyta – Nitzschia sp.). В описываемом опыте оценивали влияние отношения N/P и на размерную структуру сообщества. Оказалось, что с ростом величины N/P растёт биомасса классов мелких клеток и убывает — крупных (естественно, что меняются не размеры клеток, а доля в сообществе видов, имеющих соответствующие размеры).
Исследования in situ вели (Levich, Bulgakov, 1992; Левич и др. 1996 а,б) в течение 3-х лет на семи экспериментальных рыбоводных прудах от 0,25 до 3,5 га и глубиной около 1,5 м. В опытные и контрольные пруды с начала апреля вносили минеральные азот и фосфор в различных пропорциях: N/P = 4 в контрольных прудах и N/P = 25-50 – в опытных. Кроме того, привлекали данные о фитопланктоне и гидрохимических характеристиках тех же прудов в другие сезоны (всего около 20 “годопрудов”). Исследовали биомассы популяций фитопланктона, контролировали гидрохимические показатели прудов. Высокие отношения N/P приводили к увеличению доли протококковых водорослей и снижению доли синезелёных (в составе которых доминировали неазотфиксирующие виды).
Одним из возможных механизмов обнаруженных зависимостей численностей синезелёных и зелёных водорослей от отношения N/P может быть выявленная теоремой о максимуме видовых обилий зависимость оптимального отношения N/P от отношения соответствующих потребностей клеток (в тех случаях, если для доминирующих в ценозах видов зелёных водорослей это отношение высокое, а для синезелёных – низкое).
Подводя итог, еще раз обратим внимание читателей на то, что подход, лежащий в основе описанных теоретических и экспериментальных исследований по моделированию сообществ одноклеточных организмов, представляет собой развитие формализма Джейнса и состоит из следующих этапов.
Список литературы
501 с.