 (1)
(1)Биотехнология. 2003 (в печати)
УДК 577.4:579.841.11.017.6
Подходы к управлению составом сообщества диссоциантов Pseudomonas aeruginosa: экспериментальные данные и модельные расчеты
П.В. Фурсова, Е.С. Милько, И.А. Опарина, А.П. Левич
Международный биотехнологический центр МГУ им. М.В.Ломоносова
Каф. общей экологии, Биологический факультет, Московский государственный университет им. М.В.Ломоносова, Москва, 119992
Изучали состав и плотность смешанных культур R-, S- и M-диссоциантов бактерий Pseudomonas aeruginosa на стационарной стадии развития. Экспериментальные данные о долях диссоциантов в сообществе сравнивали с относительными численностями, полученными по расчетам в рамках вариационной модели экологического сообщества. В 36 наборах данных модельный состав поликультур отличается от опытного не более, чем на 17%, а в 29-ми из них, отклонение не превышает 12%. Относительная ошибка предсказания оптической плотности культур в 29 из 36 опытах не превышает 36%. В остальных семи случаях указаны причины более сильных отличий.
В биологических процессах с участием микроорганизмов важная роль принадлежит сообществам бактерий. Параметры процессов и урожай продуцируемых веществ в большой степени зависят от доли, которая в биомассе сообщества приходится на те или иные группы клеток. Поэтому так важны технологические приемы управления структурой сообщества, которые могут поддерживать сообщества необходимого состава, представленного группами микроорганизмов в необходимых пропорциях.
Особая ситуация в микробиологических биотехнологиях возникает из-за явления диссоциации микроорганизмов [1]: любая культура способных диссоциировать бактерий фактически является сообществом спонтанно возникающих диссоциантов. При этом активность диссоциантов, их физиологические особенности, влияющие на продуцирование биологически активных веществ, различны, что приводит к необходимости контроля за составом возникающей поликультуры: например, штаммы бактерий Pseudomonas aeruginosa входят в препарат для очистки оборудования от загрязнения тяжелыми фракциями нефти [2], причем S-диссоциант оказывается более активным в разрушении н-гекса- декана [3].
Один из приемов для формирования структуры сообществ состоит в регулировании их состава с помощью отношения компонентов минерального питания в среде [4, 5]. Для диссоциантов бактерий, например, для P.aeruginosa, также известно, что доминирование того или другого диссоцианта в почве зависит от минерального состава грунтовых вод.
По мнению авторов, экспериментальные подходы к управлению структурой сообществ должны быть предварены модельным анализом путей управления.
В настоящей работе вниманию читателей предлагается анализ роста смешанных культур диссоциантов Pseudomonas aeruginosa, а точнее – состава сообщества бактерий и его плотности на стационарной стадии развития.
Диссоцианты различаются по морфологии колоний: R-диссоциант (rough) имеет шероховатый тип колоний, S (smooth) – гладкий тип и M (mucoid) – слизистый. Одни из первых фенотипических изменений при диссоциативных переходах происходят в клеточной оболочке: капсуле, клеточной стенке, цитоплазматической мембране. Эти различия определяют многие особенности диссоциантов: изменяют скорость поступления веществ в клетки, а следовательно, и скорость роста; изменяют скорость выделения веществ из клетки; изменяют устойчивость клеток к действию физических и химических факторов; отличия в текучести мембран могут изменять активность ферментов, связанных с ними.
В настоящей работе для теоретического анализа микробиологического сообщества используется вариационная модель потребления и роста [5]. В частности, эта модель позволяет по известным потребностям физиологически различающихся групп организмов рассчитывать численности субпопуляций диссоциантов как функции ограничивающих рост ресурсов.
В отличие от методов дифференциального исчисления, которые наиболее распространенны в экологическом моделировании и позволяют описывать динамику процессов в режиме реального времени, вариационные подходы, как правило, предсказывают лишь конечное стационарное состояние сообщества [6]. Однако, на пути имитации с помощью дифференциальных уравнений возникают трудности как принципиального, так и технического характера. Принципиальная трудность состоит в том, что не существует систематических правил вывода самих уравнений. Процедуры их составления основываются на полуэмпирических закономерностях, правдоподобных рассуждениях, аналогиях и искусстве модельера. Технические трудности связаны с высокой размерностью задач по управлению структурой сообществ. Для существенно многовидовых сообществ, потребляющих многочисленные ресурсы, требуется подбор сотен коэффициентов и анализ систем из десятков уравнений. Обычные приемы снижения числа переменных – их агрегирование или учет только доминирующих групп организмов – непригодны во многих задачах экологии. С течением времени существенную роль начинают играть редкие и малочисленные виды, которые, тем самым, следует включать в число переменных на начальных этапах моделирования
Итак, на основе данных о потребностях псевдомонад в основных питательных компонентах среды в рамках вариационной модели рассчитывали относительные численности диссоциантов и общую плотность культур на стационарной стадии роста в зависимости от начального состава среды. Полученные данные сравнивали с результатами экспериментов.
МАТЕРИАЛЫ И МЕТОДЫ
Культивирование бактерий. В период с 1999 по 2001 гг. были проведены 56 опытов по выращиванию смешанных культур R-, S-, и M-диссоциантов штамма Pseudomonas aeruginosa К-2 (№ 438 в коллекции DM-MSU) (все попарные сочетания диссоциантов и поликультура из трех диссоциантов). Бактерии культивировали без пополнения запаса ресурсов на средах с различным начальным содержанием глюкозы, нитратов и фосфатов. Уровень глюкозы в средах варьировали в пределах от 0.03 до 1.6%, нитрата – от 0.010 до 0.577%, фосфат использовали на уровнях 0.005 и 0.020%.
Бактерии культивировали в пробирках на 50 мл с 10 мл среды на качалке (180 об/мин) при температуре 28°С до достижения стационарной стадии роста. В качестве посевного материала использовали односуточные культуры диссоциантов псевдомонад, выращенных на плотной среде, содержащей мясо-пептонный бульон и сусло в равных отношениях (БСА). Бактерии со скошенного агара переносили петлей в пробирку с физиологическим раствором. Плотность инокулятов каждого из диссоциантов во всех опытах выравнивали по нефелометру или по стандарту мутности до содержания клеток 107 в 1 мл. Посевной материал вносили в количестве 3%. Рост бактерий оценивали нефелометрически по плотности культуры. Соотношение диссоциантов в популяции определяли по морфологии колоний рассевом на БСА.
Расчет состава поликультур. Стационарную стадию развития культуры диссоциантов описывали решением вариационной задачи [5, 7]:
(1)
где ni – конечные искомые численности каждой из групп организмов, образующих сообщество, ![]() – количество k-го ресурса, необходимое для роста клетки группы i, в расчете на одну клетку (потребность организма группы i в ресурсе k), m – общее количество взаимонезаменимых ресурсов, потребляемых сообществом, w – число групп в сообществе, Lk – начальное содержание ресурса k в среде (Lk
– количество k-го ресурса, необходимое для роста клетки группы i, в расчете на одну клетку (потребность организма группы i в ресурсе k), m – общее количество взаимонезаменимых ресурсов, потребляемых сообществом, w – число групп в сообществе, Lk – начальное содержание ресурса k в среде (Lk![]() ).
).
Для описанной модели имеет место теорема стратификации: все пространство ресурсных факторов ![]() распадается (стратифицируется) на
распадается (стратифицируется) на ![]() непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте
непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте ![]() , где J – непустое подмножество множества ресурсов {1,2,...,m}, выполняется:
, где J – непустое подмножество множества ресурсов {1,2,...,m}, выполняется:
1) решение вариационной задачи ![]() , где
, где ![]() º
{L1, L2,..., Lm}, зависят только от тех Lk, для которых kÎ
J;
º
{L1, L2,..., Lm}, зависят только от тех Lk, для которых kÎ
J;
2) на этом решении нестрогие неравенства ![]() обращаются в строгие равенства для всех kÎ
J и в строгие неравенства для всех kÏ
J [7].
обращаются в строгие равенства для всех kÎ
J и в строгие неравенства для всех kÏ
J [7].
Теорема стратификации влечёт редукцию задачи (1) к задачам
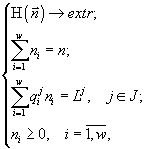 (2)
(2)
формулируемым для любого J Ì {1,2,...,m} [7].
Решение задач (2) называется формулой видовой структуры [8]
![]()
где вектора ![]() ,
, ![]() и
и ![]() имеют компоненты j из набора J, идентифицирующего страт, которому принадлежит вектор
имеют компоненты j из набора J, идентифицирующего страт, которому принадлежит вектор ![]() . Множители Лагранжа
. Множители Лагранжа ![]() и полная численность n как функции потребляемых полностью в страте
и полная численность n как функции потребляемых полностью в страте ![]() ресурсов
ресурсов ![]() получены из алгебраических уравнений
получены из алгебраических уравнений
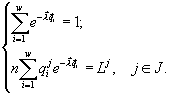
Доля диссоцианта в сообществе ![]() и полная численность клеток всего сообщества n оказываются известными функциями начальных концентраций ресурсов в среде
и полная численность клеток всего сообщества n оказываются известными функциями начальных концентраций ресурсов в среде ![]() . Знание указанной зависимости позволяет предлагать пути направленного управления долями диссоциантов в поликультуре с помощью подбора начального соотношения потребляемых клетками ресурсов. Более точно параметры управления задает теорема о максимуме видовых обилий [5, 9]. Согласно полученным результатам относительные численности видов зависят только от отношений полностью потребляемых сообществом ресурсов среды; относительная численность заданного вида принимает наибольшее значение при отношениях в среде ресурсов, равных отношениям потребностей в них данного вида.
. Знание указанной зависимости позволяет предлагать пути направленного управления долями диссоциантов в поликультуре с помощью подбора начального соотношения потребляемых клетками ресурсов. Более точно параметры управления задает теорема о максимуме видовых обилий [5, 9]. Согласно полученным результатам относительные численности видов зависят только от отношений полностью потребляемых сообществом ресурсов среды; относительная численность заданного вида принимает наибольшее значение при отношениях в среде ресурсов, равных отношениям потребностей в них данного вида.
Итак, для определения структуры сообщества на стационарной стадии роста сначала необходимо выяснить, к какому страту принадлежит вектор ресурсов, задающий состав исходной среды, а затем по формуле видовой структуры рассчитать численности групп организмов, образующих смешанную культуру.
Расчет оптической плотности культуры. Для модельного определения оптической плотности культуры бактерий на стационарной стадии роста производили следующие расчеты. Сначала находили полную численность сообщества по формуле
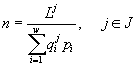 ,
,
где ![]() – начальное содержание в среде одного из полностью потребляемых сообществом ресурсов,
– начальное содержание в среде одного из полностью потребляемых сообществом ресурсов, ![]() , i = 1,...,w – относительные численности диссоциантов в смешанной культуре, отыскиваемые по формуле
, i = 1,...,w – относительные численности диссоциантов в смешанной культуре, отыскиваемые по формуле ![]() . Затем количество клеток каждого диссоцианта (соответствующая доля от общего числа клеток) переводили в оптическую плотность. Для этого использовали коэффициент, полученный при люминесцентном микроскопировании: для R-диссоцианта – 1010кл/мл·ед.неф., для S-диссоцианта – 0.3·1010 кл/мл·ед.неф., для М-диссоцианта – 0.17·1010 кл/мл·ед.неф. [10]. Общую оптическую плотность культуры получали как сумму оптических плотностей диссоциантов, образующих исследуемое сообщество.
. Затем количество клеток каждого диссоцианта (соответствующая доля от общего числа клеток) переводили в оптическую плотность. Для этого использовали коэффициент, полученный при люминесцентном микроскопировании: для R-диссоцианта – 1010кл/мл·ед.неф., для S-диссоцианта – 0.3·1010 кл/мл·ед.неф., для М-диссоцианта – 0.17·1010 кл/мл·ед.неф. [10]. Общую оптическую плотность культуры получали как сумму оптических плотностей диссоциантов, образующих исследуемое сообщество.
Относительную ошибку ![]() определения оптической плотности рассчитывали по формуле
определения оптической плотности рассчитывали по формуле ![]() , где экс. – экспериментальное значение, расч. – предсказание модели.
, где экс. – экспериментальное значение, расч. – предсказание модели.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Четыре типа смешанных культур диссоциантов P.aeruginosa выращивали на 14-ти средах (табл.1). В процессе роста измеряли оптическую плотность (в начале опыта, через 18-22 часа, затем каждые 4-8 часов до наступления стационарной стадии). В предполагаемый момент остановки роста делали рассев для определения состава культуры. Однако не во всех экспериментах удалось выполнить условие достижения стационарной стадии, поэтому часть опытных данных исключили из дальнейшего рассмотрения. Кроме того, в некоторых экспериментах со смесью двух диссоциантов к концу опыта происходило ее замещение поликультурой из трех диссоциантов, вызванное появлением третьего в результате диссоциации и последующей конкуренцией за ресурсы. Этот факт послужил причиной рассматривать указанные опыты как эксперименты со смесью трех диссоциантов.
Для модельных расчетов использовали определенные ранее значения потребностей R-, S- и M-диссоциантов в углероде, азоте и фосфоре (табл.2) [10].
В соответствии с разработанными способами нахождения границ областей лимитирования [11], для каждой опытной среды были определены ресурсы, ограничивающие рост сообщества. Оказалось, что все векторы ресурсов, определяемые начальным составом среды, принадлежат стратам с однофакторным лимитированием (полученные результаты хорошо согласуются с экспериментальными данными [12]). В области лимитирования одним ресурсом Lk, k = 1,2,3 относительные численности на стационарной стадии роста находятся из решения вариационной задачи:
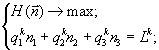
и задаются выражениями ![]() где
где ![]() – корень уравнения
– корень уравнения ![]() . Аналогично, для культуры, состоящий из двух групп организмов,
. Аналогично, для культуры, состоящий из двух групп организмов, ![]() , причем
, причем ![]() [11].
[11].
Полученные по модельным расчетам относительные численности диссоциантов сравнивали с данными экспериментов. Результаты представлены в табл.3 (номера сред соответствуют номерам табл.1). Здесь необходимо сделать существенное замечание о сопоставлении теоретических и опытных данных. Из эксперимента по результатам рассева известны доли колониеобразующих единиц (КОЕ) диссоциантов, в то время как модель позволяет рассчитывать относительные численности клеток. В рамках данной работы принята следующая гипотеза: на стационарной стадии роста клетки бактерий находятся в одинаковом физиологическом состоянии и доля проросших клеток (КОЕ) среди всех клеток данного диссоцианта одинакова для всех диссоциантов.
Таким образом, из 36 наборов данных в 29 предсказанный моделью состав поликультур отличается от экспериментального не более, чем на 12 %. Наибольшее отклонение расчетных значений от опытных составляет 17%. Согласно проведенному анализу чувствительности модели к вариациям параметров [13], полученные различия могут быть объяснены ошибками в значениях потребностей, применявшихся в расчетах (Табл.2).
В указанных 36-х опытах произвели модельный расчет общей оптической плотности культуры на стационарной стадии роста. В табл.4 приведены экспериментальные данные об оптической плотности, расчетные значения, уровень кислотности среды в тот момент времени, когда была зафиксирована максимальная оптическая плотность, относительная ошибка, а также указан лимитирующий ресурс (начальное содержание в среде именно этого ресурса используется при расчете полной численности). Номера сред соответствуют номерам из табл.1. За пределы допустимых значений ошибок, связанных с неточностью определения уровня вещества в среде и данных о потребностях, выходят данные на средах № 6 (культура SM), 10, 12 и 14. Эти несоответствия могут быть объяснены следующими образом. В среде № 6 содержалось, например, в четыре раза больше углерода, чем в среде №5, поэтому при полном исчерпании этого ресурса на обеих средах достигаемые оптические плотности должны различаться также в четыре раза. Однако, на среде № 6 измеренный уровень плотности оказался ниже, что позволяет сделать вывод о том, что стационарная стадия в опыте не была зафиксирована. Аналогично, на среде № 10 при полном исчерпании ресурса уровень оптической плотности должен быть выше (исходя из данных для среды № 9, где содержание углерода в четыре раза ниже, плотность должна достигнуть значения 272 единицы, вместо экспериментальных 182). На среде № 14 остановка роста возможно была связана не с исчерпанием питательного вещества, а с закислением среды (уровень pH равен 3.4), поэтому культура не достигла возможного уровня оптической плотности при данном содержании азота в среде (142 единицы вместо 240 единиц, которые могли быть достигнуты исходя из данных для среды № 13, где содержание азота в четыре раза ниже). На среде № 12 культура также не достигла своего максимально возможного уровня оптической плотности (215-225 единиц вместо 388-440, по данным для среды №11, где содержание фосфора в четыре раза ниже). То есть остановка роста произошла по причинам, не связанным с исчерпанием питательных веществ из среды, что подтверждается отсутствием возобновления роста после внесения ресурсов в среду (подробнее опыты с добавками описаны в статье, специально посвященной выявлению ресурсов, лимитирующих рост культур [12]). Проведенный анализ показывает, что вариационная модель адекватно предсказывает оптическую плотность смешанных культур, а некоторые расхождения экспериментальных и расчетных данных связаны с вышеописанными причинами.
Авторы выражают глубокую благодарность В.Н.Максимову и Н.Г.Булгакову за плодотворные обсуждения материалов и помощь в работе.
Работа поддержана Российским фондом фундаментальных исследований (гранты 02-04-48085 и 03-04-06044).
СПИСОК ЛИТЕРАТУРЫ
Таблица 1
Состав сред (в расчете на углерод, азот и фосфор, мг/мл) и соотношение диссоциантов (в процентах) в культуре в начале опыта
|
номер среды |
углерод |
азот |
фосфор |
R:S |
R:M |
S:M |
R:S:M |
|
1 |
0.4 |
0.035 |
0.01 |
52:48 |
61:39 |
63:37 |
37:42:21 |
|
2 |
1.62 |
0.14 |
0.04 |
52:48 |
58:42 |
51:49 |
23:53:24 |
|
3 |
0.4 |
0.015 |
0.01 |
47:53 |
59:41 |
71:29 |
35:42:23 |
|
4 |
1.6 |
0.06 |
0.04 |
47:53 |
49:51 |
54:46 |
27:34:39 |
|
5 |
0.12 |
0.035 |
0.01 |
52:48 |
61:39 |
63:37 |
37:42:21 |
|
6 |
0.48 |
0.14 |
0.04 |
52:48 |
58:42 |
51:49 |
23:53:24 |
|
7 |
0.12 |
0.015 |
0.01 |
47:53 |
59:41 |
71:29 |
35:42:23 |
|
8 |
0.48 |
0.06 |
0.04 |
47:53 |
49:51 |
54:46 |
27:34:39 |
|
9 |
0.78 |
0.1 |
0.01 |
43:57 |
49:51 |
54:46 |
29:45:26 |
|
10 |
3.24 |
0.4 |
0.04 |
43:57 |
60:40 |
55:45 |
32:42:26 |
|
11 |
1.6 |
0.2 |
0.01 |
42:58 |
59:41 |
57:43 |
43:34:23 |
|
12 |
6 |
0.8 |
0.04 |
43:57 |
60:40 |
55:45 |
32:42:26 |
|
13 |
1.2 |
0.035 |
0.01 |
43:57 |
49:51 |
54:46 |
29:45:26 |
|
14 |
4.8 |
0.14 |
0.04 |
42:58 |
59:41 |
57:43 |
43:34:23 |
Таблица 2
Значения потребностей в ресурсах среды (10-12мг/кл) и их погрешности для диссоциантов P.aeruginosa [10]
|
Диссоциант |
Ресурс |
||
|
углерод |
азот |
фосфор |
|
|
R |
129 ± 22 |
7.0 ± 2.5 |
1.0 ± 0.5 |
|
S |
409 ± 55 |
17.5 ± 3.0 |
4.0 ± 1.5 |
|
M |
525 ± 114 |
31 ± 3 |
6.0 ± 2.5 |
Таблица 3
Состав смешанных культур на стационарной стадии роста
|
номер среды из табл.1 |
2 |
3 |
5 |
||||||||||||||||||||
|
культура |
R |
S |
S |
M |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
S |
S |
M |
R |
S |
M |
R |
S |
M |
|
состав по результатам эксперимента (в процентах) |
67 |
33 |
69 |
31 |
62 |
38 |
71 |
29 |
50 |
50 |
43 |
41 |
16 |
70 |
30 |
50 |
50 |
67 |
11 |
22 |
43 |
41 |
16 |
|
состав по модельным расчетам (в процентах) |
66 |
34 |
54 |
46 |
65 |
35 |
74 |
26 |
60 |
40 |
60 |
29 |
11 |
69 |
31 |
54 |
46 |
60 |
29 |
11 |
60 |
29 |
11 |
|
номер среды из табл.1 |
6 |
7 |
8 |
|||||||||||||||||
|
культура |
R |
M |
S |
M |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
M |
S |
M |
R |
S |
M |
|
состав по результатам эксперимента (в процентах) |
60 |
40 |
42 |
58 |
61 |
39 |
62 |
38 |
50 |
50 |
62 |
32 |
6 |
78 |
22 |
43 |
57 |
68 |
17 |
15 |
|
состав по модельным расчетам (в процентах) |
73 |
27 |
54 |
46 |
69 |
31 |
73 |
27 |
54 |
46 |
62 |
23 |
15 |
73 |
27 |
54 |
46 |
62 |
23 |
15 |
|
номер среды из табл.1 |
9 |
10 |
11 |
||||||||||||||||||
|
культура |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
S |
M |
R |
S |
R |
M |
S |
M |
R |
S |
M |
|
состав по результатам эксперимента (в процентах) |
79 |
21 |
87 |
13 |
52 |
48 |
75 |
12 |
13 |
68 |
19 |
13 |
60 |
40 |
86 |
14 |
65 |
35 |
60 |
27 |
13 |
|
состав по модельным расчетам (в процентах) |
69 |
31 |
73 |
27 |
54 |
46 |
62 |
23 |
15 |
62 |
23 |
15 |
72 |
28 |
78 |
22 |
57 |
43 |
68 |
22 |
10 |
|
номер среды из табл.1 |
12 |
13 |
14 |
|||||||||||||||
|
культура |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
S |
R |
M |
R |
S |
M |
R |
M |
|
состав по результатам эксперимента (в процентах) |
62 |
38 |
84 |
16 |
72 |
28 |
63 |
22 |
15 |
70 |
30 |
80 |
20 |
45 |
31 |
24 |
75 |
25 |
|
состав по модельным расчетам (в процентах) |
72 |
28 |
78 |
22 |
57 |
43 |
68 |
22 |
10 |
65 |
35 |
74 |
26 |
60 |
29 |
11 |
74 |
26 |
Таблица 4
Оптическая плотность смешанных культур на стационарной стадии роста (в единицах нефелометра, умноженных на 100)
|
номер среды из табл. 1 |
лимитирующий ресурс |
культура |
pH |
оптическая плотность в эксперименте |
расчетная оптическая плотность |
относительная ошибка |
|
2 |
С |
RS |
6.9 |
112 |
128 |
14 |
|
SM |
7.8 |
142 |
158 |
11 |
||
|
3 |
N |
RS |
7.5 |
24 |
25.4 |
6 |
|
RM |
7.2 |
25 |
25 |
0 |
||
|
SM |
7.5 |
26 |
28.7 |
10 |
||
|
RSM |
7.4 |
25 |
26.6 |
6 |
||
|
5 |
C |
RS |
7.4 |
12 |
9.7 |
20 |
|
SM |
7.3 |
10 |
11.7 |
17 |
||
|
RSM |
7.2 |
14 |
10.7 |
24 |
||
|
RSM |
7.5 |
10 |
10.7 |
7 |
||
|
6 |
C |
RM |
7.3 |
37 |
46.4 |
25 |
|
SM |
7 |
29 |
45 |
55 |
||
|
7 |
C |
RS |
7.8 |
15 |
9.6 |
36 |
|
RM |
7.8 |
15 |
11.8 |
21 |
||
|
SM |
7.8 |
14 |
11.7 |
17 |
||
|
RSM |
8.1 |
16 |
10.6 |
34 |
||
|
8 |
C |
RM |
6.8 |
63 |
46.4 |
26 |
|
SM |
7.6-8.7 |
41 |
45 |
10 |
||
|
RSM |
7.5 |
47 |
43.1 |
8 |
||
|
9 |
C |
RS |
8.7 |
63 |
63.8 |
1 |
|
RM |
9 |
58 |
73 |
26 |
||
|
SM |
8.6 |
78 |
77.9 |
0.1 |
||
|
RSM |
8.8 |
68 |
71.7 |
5 |
||
|
10 |
C |
RSM |
7.9 |
182 |
287.2 |
58 |
|
11 |
P |
RS |
8.6 |
110 |
90 |
18 |
|
RM |
8.7 |
101 |
99.4 |
2 |
||
|
SM |
7.2 |
97 |
93 |
4 |
||
|
RSM |
8.7 |
97 |
92.1 |
5 |
||
|
12 |
P |
RS |
8.6 |
215 |
358.4 |
67 |
|
RM |
8.9 |
225 |
395 |
76 |
||
|
SM |
7.7 |
225 |
372.1 |
65 |
||
|
RSM |
8.4 |
225 |
366.1 |
63 |
||
|
13 |
N |
RS |
4.6 |
54 |
60 |
11 |
|
RM |
5.3 |
60 |
59 |
2 |
||
|
RSM |
4 |
74 |
62 |
16 |
||
|
14 |
N |
RM |
3.4 |
142 |
239.9 |
69 |