 (1)
(1)УДК 57.02.001.57
Биофизика. 2004. Т.49. № 5. С. 912-919.
© П.В.Фурсова, А.П.Левич
О РЕГУЛИРОВАНИИ СОСТАВА ЭКОЛОГИЧЕСКОГО СООБЩЕСТВА С ПОМОЩЬЮ ИЗМЕНЕНИЯ ОТНОШЕНИЯ КОНЦЕНТРАЦИЙ РЕСУРСОВ В СРЕДЕ
П.В. Фурсова, А.П. Левич
Биологический факультет МГУ им. М.В.Ломоносова, 119992, Москва
Изучена возможность регулирования состава сообщества, которую предоставляет вариационная модель экологического сообщества. Для R-, S- и M- диссоциантов бактерий Pseudomonas aeruginosa получены зависимости относительных численностей микроорганизмов от начальных отношений запасов питательных веществ в среде. Показана теоретическая возможность управления составом сообщества. Смена доминирующего вида при изменении отношения ресурсов, как по модельным расчетам, так и в эксперименте, проиллюстрирована на примере сообщества протококковых водорослей.
Ключевые слова: состав сообщества, регулирование состава сообщества, вариационная модель, Pseudomonas aeruginosa.
Одной из важнейших целей количественной экологии сообществ является умение рассчитывать численность каждой из входящих в сообщество популяций организмов как функцию доступных ресурсов среды. Практическое применение подобных расчетов заключается в возможности управлять структурой сообщества с помощью внешних факторов – изменять видовой состав и доминирование видов, целенаправленно увеличивать или уменьшать численности выбранных групп организмов.
В настоящей работе продемонстрированы возможности регулирования состава смешанной культуры микроорганизмов методом, предсказанным вариационной моделью экологического сообщества.
Вариационная модель
Предложена модель сообщества одноклеточных организмов, потребляющих ресурсы, которые не могут заменить друг друга, поскольку выполняют различные функции по отношению к росту. В модели допустимы деление и смертность клеток, но не их слияние. В лабораторных условиях описываемая модель соответствует культивированию без добавления или изъятия ресурсов и микроорганизмов. Изучено развитие поликультуры до остановки роста, вызванной исчерпанием одного из ресурсов, но не какими-либо иными причинами.
Стационарная стадия роста сообщества описана решением вариационной задачи
(1)
где ni – численности групп организмов, образующих сообщество, w – число групп в сообществе, (L1, L2,..., Lm) – количества всех потребляемых сообществом ресурсов среды в начале опыта, ![]() – количество k-ого ресурса, необходимое для деления клеток группы i, в расчете на одну клетку (потребность организма группы i в ресурсе k) [1, 2].
– количество k-ого ресурса, необходимое для деления клеток группы i, в расчете на одну клетку (потребность организма группы i в ресурсе k) [1, 2].
Целевая функция ![]() для допустимых в модели преобразований рассчитана на основе теоретико-категорной модели [3].
для допустимых в модели преобразований рассчитана на основе теоретико-категорной модели [3].
Аналитические результаты исследования задачи в зависимости от начального содержания ресурсов в среде позволяют указывать те, которые будут полностью потреблены, и рассчитывать доли каждого вида и общую численность сообщества на стационарной стадии роста.
Выявление лимитирующих (полностью потребленных) ресурсов проведено на основе теоремы стратификации [1]. Согласно этой теореме все пространство ресурсных факторов ![]() распадается (стратифицируется) на 2m-
1 непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте
распадается (стратифицируется) на 2m-
1 непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте ![]() , где J ¹
Æ
— подмножество множества ресурсов {1,2,...,m}, выполнено:
, где J ¹
Æ
— подмножество множества ресурсов {1,2,...,m}, выполнено:
Теорема стратификации влечёт редукцию задачи (1) к задачам:
 (2)
(2)
формулируемым для любого J Ì {1,2,...,m}.
Решение задач (2) называется формулой видовой структуры и имеет вид:
![]()
где ![]() , "вектор"
, "вектор" ![]() имеет компоненты
имеет компоненты ![]() с индексами из набора J, идентифицирующего страт, которому принадлежит вектор
с индексами из набора J, идентифицирующего страт, которому принадлежит вектор ![]() . Множители Лагранжа
. Множители Лагранжа ![]() и полная численность n как функции потребляемых полностью в страте
и полная численность n как функции потребляемых полностью в страте ![]() ресурсов
ресурсов ![]() ищутся из алгебраических уравнений
ищутся из алгебраических уравнений

Таким образом, в рамках вариационной модели экологического сообщества для определения его состава на стационарной стадии роста сначала необходимо выяснить, к какому страту принадлежит вектор ресурсов, задающий состав исходной среды, а затем по формуле видовой структуры рассчитать численности групп организмов, образующих сообщество.
Возможность управления составом сообщества предоставляет следующая особенность решения вариационной задачи. Установлено, что относительные численности видов на стационарной стадии роста зависят только от отношений начальных концентраций полностью потребляемых сообществом ресурсов среды [4]. Именно это свойство позволяет регулировать состав сообщества: изменяя отношения запасов питательных веществ в начальной среде, получать различные относительные численности видов на стационарной стадии роста.
Состав сообщества на стационарной стадии роста
Для демонстрации указанных возможностей, предлагается приложение вариационной модели к описанию развития сообщества R-, S-, и M-диссоциантов штамма Pseudomonas aeruginosa К-2. В период с 1999 по 2001 гг. на кафедрах микробиологии и общей экологии Биологического факультета МГУ им. М.В.Ломоносова были проведены опыты по выращиванию моно- и смешанных культур (все попарные сочетания диссоциантов и поликультура из трех диссоциантов). Частота диссоциативных переходов бактерий составляет 10-2-10-4 на одно клеточное деление, что позволяет считать сообщество диссоциантов удовлетворяющим предпосылкам вариационной модели (т.е. пренебречь явлением диссоциации в сравнении с делением). Бактерии культивировали без пополнения запаса ресурсов на средах с различным начальным содержанием глюкозы, нитратов и фосфатов.
По данным опытов с монокультурами рассчитали потребности диссоциантов в питательных веществах (табл.1) [5]. Затем, используя полученные значения, по разработанным алгоритмам [1, 6] построили области лимитирования. Для проверки адекватности предсказанной стратификации [7] применяли метод добавок. В момент предполагаемого достижения стационарной стадии культуру разделяли на четыре пробирки. В три из них вносили добавки – глюкозу, нитрат или фосфат, – четвертую оставляли без изменений (контроль). Культуру оставляли расти еще четыре часа, затем проводили измерение оптической плотности. Если добавление компонента питания в среду приводило к возобновлению деления клеток, то этот ресурс считали лимитирующим. В случае, если внесение вещества не приводило к росту культуры, считали, что этот фактор не ограничивал развития сообщества. Согласно полученным результатам (табл.2), в 75% случаев возобновление деления клеток происходило после внесения питательного вещества, предсказанного моделью как лимитирующего [7]. Возможной причиной отсутствия влияния добавок на рост бактерий является недостаточный четырехчасовой срок экспозиции культур с добавками.
Стратификация пространства ресурсов при фиксированном уровне фосфора в начальной среде для смешанной культуры трех и двух (RS) диссоциантов представлена на рис.1а и 1б (для культуры из двух диссоциантов областью лимитирования трех ресурсов является отрезок на границе IV, V и VI областей – см. рис.1б). Вид областей лимитирования в координатах ![]() для смеси двух диссоциантов изображен на рис.2.
для смеси двух диссоциантов изображен на рис.2.
В каждом из стратов, используя формулу видовой структуры, рассчитали относительные численности групп организмов, образующих сообщество (вывод соответствующих формул содержится в одной из предшествующих работ [6]). В работе полностью представлен случай R- и S-диссоциантов в смеси. Зависимость относительных численностей видов в сообществе от отношения ресурсов в среде можно проследить по рис.3. Каждый из рис. 3а, 3б, 3в, 3г (все возможные конфигурации состава культуры в зависимости от соотношений питательных веществ в начале культивирования) соответствует фиксированному отношению C/P. В областях I, II, III относительные численности постоянны. Численные значения задаются выражениями ![]()
![]() где
где ![]() – корень уравнения
– корень уравнения ![]() , r и s – относительные численности R- и S-диссоциантов, индекс k обозначает ресурс, а параметры
, r и s – относительные численности R- и S-диссоциантов, индекс k обозначает ресурс, а параметры ![]() – соответствующие потребности (значения из табл.1). В области IV графиками зависимостей относительных численностей от отношения C/N являются участки гипербол, задаваемых уравнениями:
– соответствующие потребности (значения из табл.1). В области IV графиками зависимостей относительных численностей от отношения C/N являются участки гипербол, задаваемых уравнениями: ![]() ,
, ![]() , где
, где ![]() . В области ограничения роста азотом и фосфором (область VI) решение соответствующей редуцированной задачи (2) задается формулами
. В области ограничения роста азотом и фосфором (область VI) решение соответствующей редуцированной задачи (2) задается формулами ![]() ,
, ![]() , где
, где ![]() . Для построения графиков этих зависимостей на плоскости (C/N; отн.числ.) (рис.3) значения
. Для построения графиков этих зависимостей на плоскости (C/N; отн.числ.) (рис.3) значения ![]() , соответствующее области VI, рассчитывается исходя из заданного уровня отношения
, соответствующее области VI, рассчитывается исходя из заданного уровня отношения ![]() и интервала значений, принимаемых отношением
и интервала значений, принимаемых отношением ![]() , а именно, как частное
, а именно, как частное ![]() . В области V постоянные относительные численности при фиксированном отношении
. В области V постоянные относительные численности при фиксированном отношении ![]() определяются по формулам
определяются по формулам ![]() ,
, ![]() , подстановкой в них конкретного значения
, подстановкой в них конкретного значения ![]() .
.
В серии экспериментов, проведенных со смешанными культурами бактерий P.aeruginosa среды были ограничены по одному из питательных веществ. В соответствующих областях однофакторного лимитирования предсказываемый моделью состав культуры сравнивали с данными эксперимента [8]. Анализ результатов (табл.3) показывает, что из 36 наборов данных в 29 расчетный состав поликультур отличается от экспериментального не более, чем на 12 %. Максимальное отклонение предсказанных моделью значений от опытных составляет 17%. Полученные различия могут быть объяснены ошибками в значениях потребностей, применявшихся в расчетах (табл.1), о чем свидетельствует проведенный анализ чувствительности модели к вариациям параметров [9]. Провести опытную проверку предсказываемых моделью значений во всем диапазоне изменений отношений ресурсов не представляется возможным. Это связано, во-первых, с узостью интервалов Lk/Ls (k, s = 1, 2, 3), задающих области многофакторного лимитирования (см. рис.2 и рис.3), и с точностью, с которой могут быть составлены исходные среды, а во-вторых, с незначительными различиями состава сообщества в стратах в сравнении с погрешностью определения долей диссоциантов.
Регулирование состава возможно лишь в задаваемых моделью пределах. Как видно из рис.3, для видов, обладающих полученными значениями потребностей, нельзя изменить доминирующую группу организмов. Однако, при "подходящих" значениях потребностей, варьируя соотношение питательных веществ в среде, на стационарной стадии роста можно получать различные распределения видов по рангам (от наименее до наиболее многочисленного). В качестве демонстрации указанного свойства приведем расчеты состава сообщества, состоящего из четырех протококковых водорослей, потребляющих азот (N) и фосфор (P) [10]. На рис.4 представлены относительные численности видов ![]() , исходя из следующих экспериментальных значений потребностей (размерность 10-9 мг/кл):
, исходя из следующих экспериментальных значений потребностей (размерность 10-9 мг/кл):
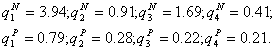
По результатам лабораторных экспериментов смену доминирующего вида можно проследить на примере культивирования двух видов из порядка протококковых Ankistrodesmus falcatus и Scenedesmus quadricauda [10, 11]. Результаты исследования роста сообщества на средах с различными начальными отношениями азота и фосфора (N/P = 1.3; 57) проиллюстрированы на рис.5.
В заключении отметим еще одно свойство вариационной задачи, имеющее отношение к управлению (или регулированию) составом сообщества: относительная численность заданного вида принимает наибольшее значение при отношениях в среде ресурсов, равных отношениям потребностей в них заданного вида [4]. Данные экспериментов подтверждают этот теоретический факт. Так, например, полученное из опытов отношение потребностей в азоте и фосфоре для A. falcatus равно 7, а для S. quadricauda – 25. Следовательно, на средах с отношениями ресурсов в среде 1.3 и 57 должен преобладать анкистродесмум и сценедесмус, соответственно. Именно это и наблюдалось в эксперименте (рис.5) (Более подробное описание указанного опыта, а также других экспериментов содержится в книге, посвященной управлению структурой сообществ фитопланктона [10]). Таким образом, сформулированное свойство относительных численностей сообщества также может быть использовано при целенаправленном влиянии на структуру сообщества.
Работа поддержана Российским фондом фундаментальных исследований (гранты 02-04-48085 и 03-04-06044).
СПИСОК ЛИТЕРАТУРЫ
Таблица 1. Значения потребностей в ресурсах среды (10-12мг/кл) и их погрешности для диссоциантов P.aeruginosa [5]
|
Диссоциант |
Ресурс |
||
|
углерод |
азот |
фосфор |
|
|
R |
129 ± 22 |
7.0 ± 2.5 |
1.0 ± 0.5 |
|
S |
409 ± 55 |
17.5 ± 3.0 |
4.0 ± 1.5 |
|
M |
525 ± 114 |
31 ± 3 |
6.0 ± 2.5 |
Таблица 2. Влияние добавок на оптическую плотность моно- и смешанных культур [7]
|
номер опыта |
лимитирующий ресурс по модели |
добавка |
R |
S |
M |
RS |
RM |
SM |
RSM |
|
C |
121 |
220 |
110 |
||||||
|
1 |
С |
N |
91 |
102 |
96 |
||||
|
P |
62 |
100 |
94 |
||||||
|
C |
101 |
||||||||
|
2 |
N |
N |
131 |
||||||
|
P |
100 |
||||||||
|
C |
117 |
||||||||
|
3 |
P |
N |
119 |
||||||
|
P |
125 |
||||||||
|
C |
220 |
140 |
217 |
||||||
|
4 |
C |
N |
97 |
80 |
74 |
||||
|
P |
106 |
115 |
109 |
||||||
|
C |
227 |
||||||||
|
5 |
C |
N |
73 |
||||||
|
P |
100 |
||||||||
|
C |
93 |
||||||||
|
6 |
N |
N |
93 |
||||||
|
P |
98 |
||||||||
|
C |
94 |
62 |
100 |
||||||
|
7 |
P |
N |
95 |
100 |
68 |
||||
|
P |
134 |
114 |
105 |
||||||
|
C |
82 |
89 |
87 |
||||||
|
8 |
C |
N |
59 |
84 |
98 |
||||
|
P |
100 |
78 |
81 |
||||||
|
C |
95 |
100 |
90 |
92 |
96 |
104 |
96 |
||
|
9 |
N |
N |
143 |
112 |
100 |
104 |
104 |
116 |
128 |
|
P |
95 |
104 |
97 |
96 |
92 |
100 |
100 |
||
|
C |
122 |
160 |
142 |
160 |
160 |
150 |
|||
|
10 |
C |
N |
100 |
90 |
75 |
100 |
90 |
71 |
|
|
P |
100 |
110 |
75 |
100 |
90 |
71 |
|||
|
C |
212 |
97 |
121 |
||||||
|
11 |
C |
N |
162 |
66 |
79 |
||||
|
P |
235 |
89 |
83 |
||||||
|
C |
129 |
129 |
143 |
120 |
120 |
113 |
|||
|
12 |
C |
N |
86 |
100 |
86 |
100 |
93 |
75 |
|
|
P |
86 |
93 |
100 |
80 |
87 |
100 |
|||
|
C |
131 |
101 |
119 |
103 |
143 |
||||
|
13 |
C |
N |
76 |
68 |
81 |
81 |
102 |
||
|
P |
87 |
57 |
72 |
73 |
92 |
||||
|
C |
112 |
103 |
109 |
129 |
|||||
|
14 |
P |
N |
91 |
88 |
81 |
91 |
|||
|
P |
108 |
94 |
95 |
82 |
Таблица 3. Состав смешанных культур на стационарной стадии роста [8]
|
номер |
1 |
2 |
3 |
||||||||||||||||||||
|
культура |
R |
S |
S |
M |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
S |
S |
M |
R |
S |
M |
R |
S |
M |
|
состав по результатам эксперимента (в процентах) |
67 |
33 |
69 |
31 |
62 |
38 |
71 |
29 |
50 |
50 |
43 |
41 |
16 |
70 |
30 |
50 |
50 |
67 |
11 |
22 |
43 |
41 |
16 |
|
состав по модельным расчетам (в процентах) |
66 |
34 |
54 |
46 |
65 |
35 |
74 |
26 |
60 |
40 |
60 |
29 |
11 |
69 |
31 |
54 |
46 |
60 |
29 |
11 |
60 |
29 |
11 |
|
номер |
4 |
5 |
6 |
|||||||||||||||||
|
культура |
R |
M |
S |
M |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
M |
S |
M |
R |
S |
M |
|
состав по результатам эксперимента (в процентах) |
60 |
40 |
42 |
58 |
61 |
39 |
62 |
38 |
50 |
50 |
62 |
32 |
6 |
78 |
22 |
43 |
57 |
68 |
17 |
15 |
|
состав по модельным расчетам (в процентах) |
73 |
27 |
54 |
46 |
69 |
31 |
73 |
27 |
54 |
46 |
62 |
23 |
15 |
73 |
27 |
54 |
46 |
62 |
23 |
15 |
|
номер |
7 |
8 |
9 |
||||||||||||||||||
|
культура |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
S |
M |
R |
S |
R |
M |
S |
M |
R |
S |
M |
|
состав по результатам эксперимента (в процентах) |
79 |
21 |
87 |
13 |
52 |
48 |
75 |
12 |
13 |
68 |
19 |
13 |
60 |
40 |
86 |
14 |
65 |
35 |
60 |
27 |
13 |
|
состав по модельным расчетам (в процентах) |
69 |
31 |
73 |
27 |
54 |
46 |
62 |
23 |
15 |
62 |
23 |
15 |
72 |
28 |
78 |
22 |
57 |
43 |
68 |
22 |
10 |
|
номер |
10 |
11 |
12 |
|||||||||||||||
|
культура |
R |
S |
R |
M |
S |
M |
R |
S |
M |
R |
S |
R |
M |
R |
S |
M |
R |
M |
|
состав по результатам эксперимента (в процентах) |
62 |
38 |
84 |
16 |
72 |
28 |
63 |
22 |
15 |
70 |
30 |
80 |
20 |
45 |
31 |
24 |
75 |
25 |
|
состав по модельным расчетам (в процентах) |
72 |
28 |
78 |
22 |
57 |
43 |
68 |
22 |
10 |
65 |
35 |
74 |
26 |
60 |
29 |
11 |
74 |
26 |
а)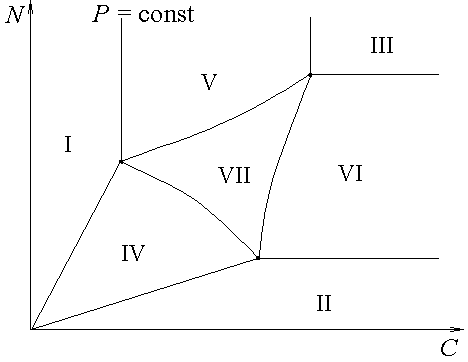
б)
Рис.1. Стратификация пространства ресурсных факторов для смешанной культуры трех (а) и двух (б) диссоциантов. В области I лимитирует углерод (С), в II – азот (N), в III – фосфор (P), в IV – углерод и азот, в V – углерод и фосфор, в VI – азот и фосфор, в VII – все три ресурса.

Рис.2. Стратификация пространства ресурсов в координатах ![]() для смеси двух диссоциантов (RS). Горизонтальные пунктирные линии соответствуют значениям отношения C/P, для которых проведены расчеты зависимости относительных обилий диссоциантов как функций отношения C/N (см. рис.3).
для смеси двух диссоциантов (RS). Горизонтальные пунктирные линии соответствуют значениям отношения C/P, для которых проведены расчеты зависимости относительных обилий диссоциантов как функций отношения C/N (см. рис.3).
г)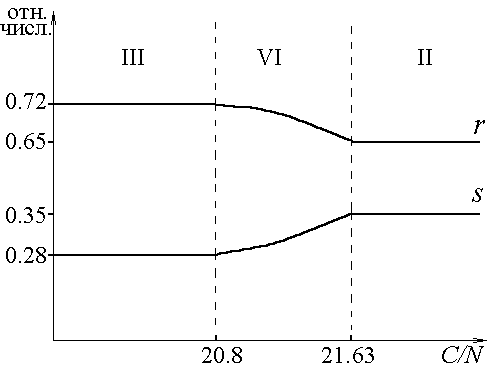
в)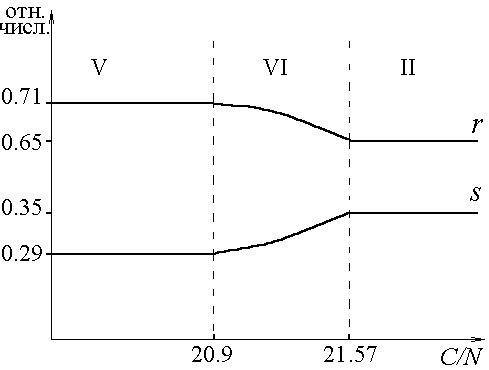
б)
а)
Рис.3. Зависимость относительных численностей (отн.числ.) R- и S-диссоциантов (r, s) от отношения углерода к азоту (C/N) в исходной среде. а) C/P ![]() (0,110.83]; б) C/P = 111; в) C/P = 112.5; г) C/P = 112.9.
(0,110.83]; б) C/P = 111; в) C/P = 112.5; г) C/P = 112.9.
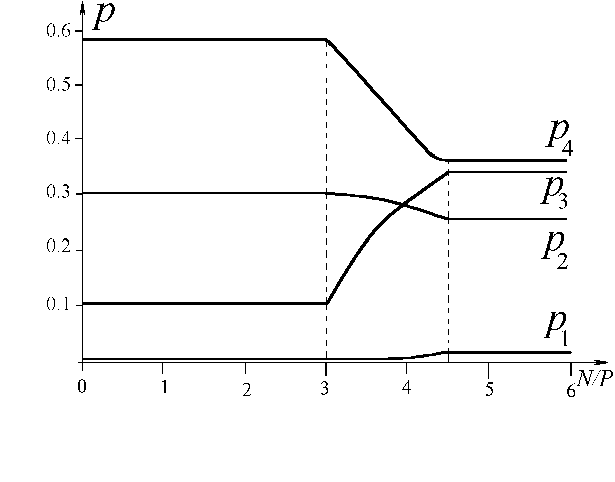
Рис.4. Относительные численности ![]() для экспериментальных потребностей как функции от N/P [10].
для экспериментальных потребностей как функции от N/P [10].

Рис.5. Зависимость относительных численностей видов от отношения азота к фосфору в лабораторных экспериментах с двухвидовыми культурами.: 1 – S. quadricauda, 2 – A. falcatus [10].
ON ECOLOGICAL COMMUNITY STRUCTURE REGULATION BY RESOURCES CONCENTRATION RATIO IN ENVIRONMENT CHANGING
P.V. Fursova, A.P. Levich
Biological faculty, MSU, 119992, Moscow
The opportunity of community structure regulation, which is given by variational model of ecological community, is investigated. For R-, S- and M- dissociants of bacteria Pseudomonas aeruginosa microorganisms relative abundances dependencies on the initial resources concentration ratio in environment are received. The theoretical opportunity of community structure control is shown. The change of a dominant species under resources concentration ratio change both on modelling accounts and in experiment is illustrated on an example of Chlorococcales community.
Key words: community structure, community structure regulation, variational model, Pseudomonas aeruginosa.