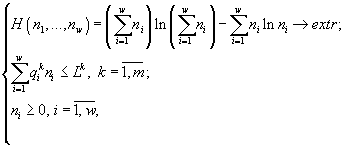
Вестник МГУ. Сер. биологическая. 2003 (в печати)
УДК 579.841.11.017.6
Выявление компонентов питания, ограничивающих рост моно- и смешанных культур диссоциантов бактерий Pseudomonas aeruginosa
П.В. Фурсова, Е.С. Милько, И.А. Опарина, А.П. Левич
(кафедра общей экологии и кафедра микробиологии)
Актуальной проблемой экологии сообществ, в частности микробиологических, является задача определения фактора, ограничивающего их рост. В настоящей работе представлена модель, в рамках которой возможно предсказание лимитирующих ресурсов, их экспериментальное определение и сравнение полученных результатов.
Материалы и методы
В период с 1999 по 2001 гг. были проведены 116 опытов по культивированию R-, S-, и M-диссоциантов штамма Pseudomonas aeruginosa К-2. Моно- и смешанные культуры бактерий культивировали без пополнения запаса ресурсов на средах с различным начальным содержанием глюкозы, нитратов и фосфатов. Уровень глюкозы в средах варьировали в пределах от 0.03 до 1.6%, нитрата – от 0.01 до 0.577%, фосфата – от 0.001 до 0.04%.
1. Методы культивирования. Бактерии культивировали в пробирках на 50 мл с 10 мл среды на качалке (180 об/мин) при температуре 28°С до достижения стационарной стадии роста. В качестве посевного материала использовали односуточные культуры диссоциантов псевдомонад, выращенных на плотной среде, содержащей мясо-пептонный бульон и сусло в отношении 1:1 (БСА). Бактерии со скошенного агара переносили петлей в пробирку с физиологическим раствором. Плотность инокулятов каждого из диссоциантов во всех опытах выравнивали по нефелометру или по стандарту мутности до содержания клеток 109 (для смешанных культур – 107) в 1 мл. Посевной материал вносили в количестве 3%. Рост бактерий оценивали нефелометрически по плотности культуры. Соотношение диссоциантов в популяции определяли по морфологии колоний рассевом на БСА.
Для экспериментального определения ресурса, ограничивающего рост культуры, проводили опыты с добавками. В момент предполагаемого достижения стационарной стадии культуру разделяли на четыре пробирки. В три из них вносили добавки – глюкозу, нитрат или фосфат, – четвертую оставляли без изменений (контроль). Культуру оставляли расти еще четыре часа, затем проводили измерение оптической плотности. Если добавление компонента питания в среду приводило к возобновлению деления клеток, то этот ресурс считали лимитирующим. В случае, если внесение вещества не приводило к росту культуры, считали, что этот фактор не ограничивал развития сообщества.
2. Метод расчета лимитирующего ресурса для поликультур. Для описания стационарной стадии развития культуры диссоциантов применяли вариационную модель, которая ранее успешно была использована при изучении структуры фитопланктонных сообществ (Левич, 1980; Левич с соавт., 1997; Levich, 2000)
где ni – конечные искомые численности каждой из групп организмов, образующих сообщество, ![]() – количество k-го ресурса, необходимое для роста клетки группы i, в расчете на одну клетку (потребность организма группы i в ресурсе k), m – общее количество взаимонезаменимых ресурсов, потребляемых сообществом, w – число групп в сообществе, Lk – начальное содержание ресурса k в среде (Lk
– количество k-го ресурса, необходимое для роста клетки группы i, в расчете на одну клетку (потребность организма группы i в ресурсе k), m – общее количество взаимонезаменимых ресурсов, потребляемых сообществом, w – число групп в сообществе, Lk – начальное содержание ресурса k в среде (Lk![]() ).
).
Для описанной модели имеет место теорема стратификации (Левич с соавт., 1994; Levich, 2000): все пространство ресурсных факторов ![]() распадается (стратифицируется) на
распадается (стратифицируется) на ![]() непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте SJ, где J – непустое подмножество множества ресурсов {1,2,...,m}, выполняется:
непересекающихся областей (стратов), каждая из которых соответствует одному из подмножеств множества потребляемых сообществом ресурсов. В страте SJ, где J – непустое подмножество множества ресурсов {1,2,...,m}, выполняется:
1) решение вариационной задачи ![]() , где
, где ![]() º
{L1, L2,..., Lm}, зависят только от тех Lk, для которых kÎ
J;
º
{L1, L2,..., Lm}, зависят только от тех Lk, для которых kÎ
J;
2) на этом решении нестрогие неравенства ![]() обращаются в строгие равенства для всех kÎ
J и в строгие неравенства для всех kÏ
J.
обращаются в строгие равенства для всех kÎ
J и в строгие неравенства для всех kÏ
J.
Биологическая интерпретация теоремы стратификации задаёт правило лимитирующего звена для многовидовых сообществ, популяции из которых конкурируют за несколько ресурсов (Левич с соавт., 1993). Лимитирующими называются ресурсы, потребляемые сообществом из среды полностью, т.е. те, для которых балансовые неравенства сформулированной задачи превращаются в равенства. Согласно теореме стратификации, из заданной совокупности ресурсов ![]() полностью потребляются, т.е. лимитируют рост сообщества, ресурсы из набора J, идентифицирующего страт SJ, которому принадлежит заданный вектор
полностью потребляются, т.е. лимитируют рост сообщества, ресурсы из набора J, идентифицирующего страт SJ, которому принадлежит заданный вектор ![]() . Таким образом, теорема стратификации позволяет строго предсказывать ресурсы, лимитирующие рост сообщества с заданным набором потребностей
. Таким образом, теорема стратификации позволяет строго предсказывать ресурсы, лимитирующие рост сообщества с заданным набором потребностей ![]() .
.
3. Метод расчета лимитирующего ресурса для монокультур. Для монокультур, т.е. в случае w = 1, целевая функция H(n1,...,nw), является тождественным нулем и численность группы определяется системой ресурсных ограничений. В ситуации, когда питательная среда содержит три компонента, имеет место система:
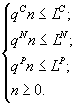
Здесь верхние индексы C, N, P обозначают, соответственно, углерод, азот и фосфор. Решение системы имеет вид: 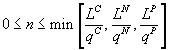 . Достижение культурой стационарной стадии означает исчерпание ресурса или ресурсов, для которого или которых реализуется указанный минимум. На математическом языке полное потребление выражается обращением соответствующего неравенства в равенство:
. Достижение культурой стационарной стадии означает исчерпание ресурса или ресурсов, для которого или которых реализуется указанный минимум. На математическом языке полное потребление выражается обращением соответствующего неравенства в равенство: ![]() , где индекс r обозначает исчерпанный ресурс. Таким образом, для аналитического выявления лимитирующего питательного компонента необходимо найти минимальное из отношений начального содержания потребляемых ресурсов и соответствующих потребностей. Возможна следующая геометрическая интерпретация. В трехмерном пространстве ресурсов вектором потребностей
, где индекс r обозначает исчерпанный ресурс. Таким образом, для аналитического выявления лимитирующего питательного компонента необходимо найти минимальное из отношений начального содержания потребляемых ресурсов и соответствующих потребностей. Возможна следующая геометрическая интерпретация. В трехмерном пространстве ресурсов вектором потребностей ![]() задается прямая. Участки трех плоскостей, проходящих через прямую q и координатные оси ресурсов, делят это пространство на 3 области лимитирования (рис.1). В каждой из этих областей рост культуры ограничен одним из трех ресурсов, а именно тем, координатная ось которого не лежит в данной области (Максимов с соавт., 2001).
задается прямая. Участки трех плоскостей, проходящих через прямую q и координатные оси ресурсов, делят это пространство на 3 области лимитирования (рис.1). В каждой из этих областей рост культуры ограничен одним из трех ресурсов, а именно тем, координатная ось которого не лежит в данной области (Максимов с соавт., 2001).
Разбиение пространства трех ресурсов на области, в которых лимитирующими являются один, два или три фактора, представлено на рис.2.
Для выявления ресурсов, ограничивающих рост смешанных культур, сначала на основе полученных ранее алгоритмов (Левич с соавт., 1994; Фурсова, 2003) рассчитывали границы областей лимитирования, а затем проверяли, к какой из них относится опытная среда.
Результаты и обсуждение
Были проведены опыты с добавками по выращиванию монокультур диссоциантов на 20-ти средах, а по выращиванию смешанных культур (попарные смеси и поликультура всех трех диссоциантов) – на 14 средах (состав сред в расчете на элемент представлен в табл.1а, б).
Для указанных сред было проведено модельное выявление лимитирующих ресурсов. В расчетах использовали ранее определенные (Фурсова с соавт., 2004) значения потребностей (табл.2).
Затем полученные данные сравнивали с результатами опытов с добавками. Необходимо отметить, что ряд экспериментов исключили из рассмотрения. Это связано с двумя обстоятельствами:
Из 116 проведенных экспериментов 47 удовлетворяли необходимым требованиям. В табл.3а,б указаны номера сред (в соответствии с табл.1а,б), предсказываемый модельными расчетами лимитирующий ресурс и процентное изменение оптической плотности культуры в результате добавления питательного вещества по отношению к контролю. В 35-ти экспериментах рост возобновился после добавки вещества, определенного моделью как лимитирующего.
Таким образом, полученные результаты позволяют сделать вывод о том, что теорема стратификации в 75% случаев дает адекватное предсказание ресурсов, ограничивающих рост микробиологического сообщества.
Возможной причиной отсутствия влияния добавок на рост бактерий является недостаточный четырехчасовой срок экспозиции культур с добавками. Чтобы избежать влияния лагфазы, в дальнейших экспериментах следует проверять наличие или отсутствие возобновления роста не через 4, а через 8-16 часов после внесения добавок. Еще одним уточнением методики дальнейших опытов с добавками была бы необходимость вносить не только отдельные ресурсы, но и их комбинации для проверки многофакторного лимитирования (т.е. по достижении стационарной стадии разделять культуру не на 4, а на 7 пробирок).
Авторы выражают глубокую благодарность В.Н.Максимову и Н.Г.Булгакову за плодотворные обсуждения материалов и помощь в работе.
***
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (грант 02-04-48085).
Список литературы
Левич А.П. 1980. Структура экологических сообществ. М.
Левич А.П., Алексеев В.Л., Никулин В.А. 1994. Математические аспекты вариационного моделирования в экологии сообществ // Математическое моделирование. 6, № 5. 55–76.
Левич А.П., Замолодчиков Д.Г., Алексеев В.Л. 1993. Правило лимитирующего звена для многовидовых экологических сообществ // Журн. общ. биологии. 54, № 3. 282–297.
Левич А.П., Максимов В.Н., Булгаков Н.Г. 1997. Теоретическая и экспериментальная экология фитопланктона. Управление структурой и функциями сообществ. Учебное пособие. М.
Максимов В.Н., Милько Е.С., Левич А.П. 2001. Потребности диссоциантов Pseudomonas aeruginosa в глюкозе, нитратах и фосфатах. Лимитирующие рост концентрации веществ при культивировании без пополнения запаса ресурсов // Изв. АН. Сер. биол. № 5. 607–612.
Фурсова П.В. 2003 (в печати). Расчеты видовых обилий и областей лимитирования в вариационной модели экологического сообщества // Математическое моделирование.
Фурсова П.В., Милько Е.С., Опарина И.А., Максимов В.Н., Левич А.П. 2004 (в печати). Определение потребностей диссоциантов Pseudomonas aeruginosa в углероде, азоте и фосфоре по результатам культивирования без пополнения запасов ресурсов // Микробиология.
Levich A.P. 2000. Variational modelling theorems and algocoenoses functioning principles // Ecological Modelling. 131. 207–227.
Таблица 1а
Состав сред по углероду, азоту и фосфору в начале опыта для монокультур (мг/мл)
|
номер среды |
R-диссоциант |
S- и M- диссоцианты |
||||
|
углерод |
азот |
фосфор |
углерод |
азот |
фосфор |
|
|
1 |
0.78 |
0.4 |
0.028 |
0.78 |
0.4 |
0.028 |
|
2 |
3.18 |
0.1 |
0.028 |
3.18 |
0.1 |
0.028 |
|
3 |
3.18 |
0.4 |
0.007 |
3.18 |
0.4 |
0.007 |
|
4 |
0.282 |
0.1 |
0.008 |
0.282 |
0.1 |
0.008 |
|
5 |
1.6 |
0.03 |
0.008 |
0.78 |
0.03 |
0.008 |
|
6 |
1.6 |
0.1 |
0.002 |
0.78 |
0.1 |
0.002 |
Таблица 1б
Состав сред по углероду, азоту и фосфору в начале опыта для моно- и смешанных культур (мг/мл)
|
номер среды |
углерод |
азот |
фосфор |
|
1 |
0.4 |
0.035 |
0.01 |
|
2 |
1.62 |
0.14 |
0.04 |
|
3 |
0.4 |
0.015 |
0.01 |
|
4 |
1.6 |
0.06 |
0.04 |
|
5 |
0.12 |
0.035 |
0.01 |
|
6 |
0.48 |
0.14 |
0.04 |
|
7 |
0.12 |
0.015 |
0.01 |
|
8 |
0.48 |
0.06 |
0.04 |
|
9 |
0.78 |
0.1 |
0.01 |
|
10 |
3.24 |
0.4 |
0.04 |
|
11 |
1.6 |
0.2 |
0.01 |
|
12 |
6 |
0.8 |
0.04 |
|
13 |
1.2 |
0.035 |
0.01 |
|
14 |
4.8 |
0.14 |
0.04 |
Таблица 2
Значения потребностей (10-12мг/кл) диссоциантов P.aeruginosa в ресурсах среды (Фурсова с соавт., 2004)
|
Диссоциант |
Ресурс |
||
|
углерод |
азот |
фосфор |
|
|
R |
129 |
7 |
1 |
|
S |
409 |
17 |
4 |
|
M |
525 |
31 |
6 |
Таблица 3а
Влияние добавок на оптическую плотность монокультур
|
номер среды |
лимитирующий ресурс по модели |
добавка |
R |
S |
M |
|
C |
121 |
220 |
110 |
||
|
1 |
С |
N |
91 |
102 |
96 |
|
P |
62 |
100 |
94 |
||
|
C |
101 |
||||
|
2 |
N |
N |
131 |
||
|
P |
100 |
||||
|
C |
117 |
||||
|
3 |
P |
N |
119 |
||
|
P |
125 |
||||
|
C |
220 |
140 |
217 |
||
|
4 |
C |
N |
97 |
80 |
74 |
|
P |
106 |
115 |
109 |
||
|
C |
227 |
||||
|
4* |
C |
N |
73 |
||
|
P |
100 |
||||
|
C |
93 |
||||
|
5 |
N |
N |
93 |
||
|
P |
98 |
||||
|
C |
94 |
62 |
100 |
||
|
6 |
P |
N |
95 |
100 |
68 |
|
P |
134 |
114 |
105 |
*Вторая повторность опыта с S-диссоциантом
Таблица 3б
Влияние добавок на оптическую плотность моно- и смешанных культур
|
номер среды |
лимитирующий ресурс по модели |
добавка |
R |
S |
M |
RS |
RM |
SM |
RSM |
|
C |
82 |
89 |
87 |
||||||
|
2 |
C |
N |
59 |
84 |
98 |
||||
|
P |
100 |
78 |
81 |
||||||
|
C |
95 |
100 |
90 |
92 |
96 |
104 |
96 |
||
|
3 |
N |
N |
143 |
112 |
100 |
104 |
104 |
116 |
128 |
|
P |
95 |
104 |
97 |
96 |
92 |
100 |
100 |
||
|
C |
122 |
160 |
142 |
160 |
160 |
150 |
|||
|
5 |
C |
N |
100 |
90 |
75 |
100 |
90 |
71 |
|
|
P |
100 |
110 |
75 |
100 |
90 |
71 |
|||
|
C |
212 |
97 |
121 |
||||||
|
6 |
C |
N |
162 |
66 |
79 |
||||
|
P |
235 |
89 |
83 |
||||||
|
C |
129 |
129 |
143 |
120 |
120 |
113 |
|||
|
7 |
C |
N |
86 |
100 |
86 |
100 |
93 |
75 |
|
|
P |
86 |
93 |
100 |
80 |
87 |
100 |
|||
|
C |
131 |
101 |
119 |
103 |
143 |
||||
|
9 |
C |
N |
76 |
68 |
81 |
81 |
102 |
||
|
P |
87 |
57 |
72 |
73 |
92 |
||||
|
C |
112 |
103 |
109 |
129 |
|||||
|
11 |
P |
N |
91 |
88 |
81 |
91 |
|||
|
P |
108 |
94 |
95 |
82 |
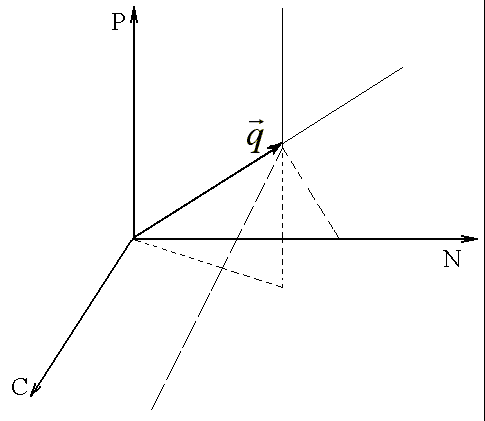
Рис. 1. Разбиение пространства ресурсов на области лимитирования роста монокультуры.
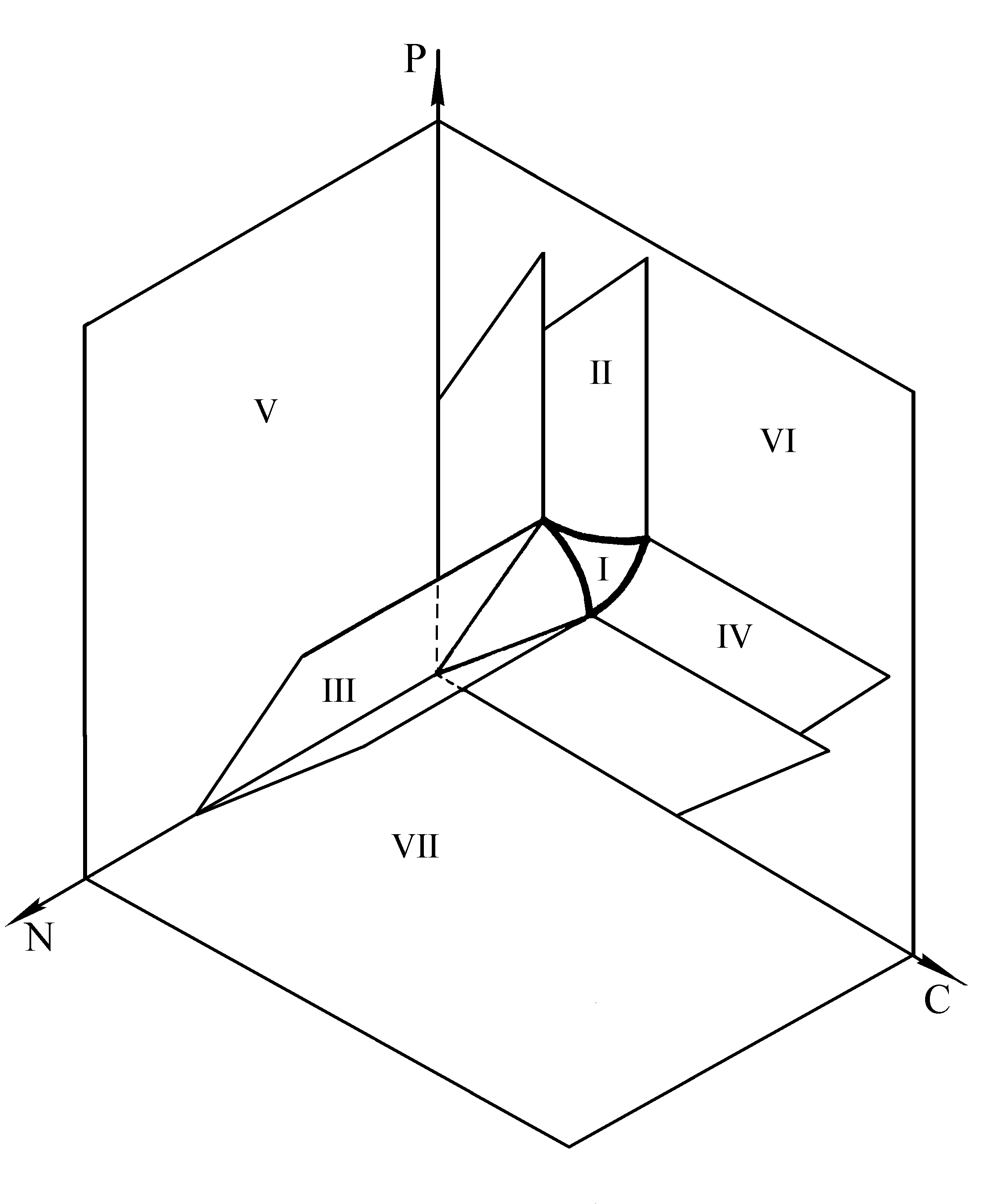
Рис. 2. Разбиение пространства ресурсов на области лимитирования: в области I лимитирующими являются три ресурса, в области II – L1 и L2; в III – L1 и L3; в IV – L2 и L3; в V – L1; в VI – L2; в VII – L3.
Revealing of the resources, limiting mono- and mixed Pseudomonas aeruginosa dissociants cultures growth
The model and experimental revealing of limiting resources on the data of experiments with mono- and mixed Pseudomonas aeruginosa dissociants cultures carried out. In 75 % of cases the theoretical calculation tally with independent experimental results. The probable explanations of results discrepancies, and also specification to methods of the subsequent experiment realization are offered.