Лаборатория общей экологии Биологического факультета Московского государственного университета им. М.В.Ломоносова (119899, Москва, Воробьевы горы; e-mail:
 )
)
Феноменология, применение и происхождение ранговых распределений в биоценозах и экологии как источник идей для техноценозов и ЭКОНОМИКИ
А.П.Левич
Лаборатория общей экологии Биологического факультета Московского государственного университета им. М.В.Ломоносова (119899, Москва, Воробьевы горы; e-mail:
)
Ранговые распределения численностей или биомасс групп живых организмов представляют собой действенный инструмент количественного исследования экологических сообществ. В качестве групп могут выступать биологические таксоны, размерные классы, совокупности особей, объединенные по каким-либо физиологическим или иным признакам. В обзорах по применению ранговых распределений в экологии (Левич, 1980; Frontier, 1985; Пузаченко, 1996) можно найти немало ссылок на исследования сообществ планктона, бентоса, перифитона, птиц, млекопитающих, насекомых, высших растений и многого другого.
Проблемы, связанные с методом, можно объединить в три большие группы: феноменология, применение и происхождение ранговых распределений.
Описание распределений
Для феноменологического описания ранговых распределений (Левич, 1980) в экологии применяются различные аппроксимации: экспоненциальная модель ![]()
![]() , гиперболическоя модель
, гиперболическоя модель ![]()
![]() , объединяющее их дзета-распределение
, объединяющее их дзета-распределение ![]() ~
~![]() , модель “разломанного стержня”
, модель “разломанного стержня” ![]() ~
~![]() (в формулах
(в формулах ![]() обозначает численность особей ранга i; z, b и B — параметры моделей).
обозначает численность особей ранга i; z, b и B — параметры моделей).
Вместо функций ![]() иногда анализируют их математические эквиваленты: распределения накопленных численностей (суммарную численность групп всех рангов от 1 до i), частотные распределения (число групп с численностями от n до n+Dn) или частотные распределения логарифмов численностей, функции экологической неаддитивности (зависимость числа групп в выборке от объема выборки).
иногда анализируют их математические эквиваленты: распределения накопленных численностей (суммарную численность групп всех рангов от 1 до i), частотные распределения (число групп с численностями от n до n+Dn) или частотные распределения логарифмов численностей, функции экологической неаддитивности (зависимость числа групп в выборке от объема выборки).
Общий вывод об адекватности различных моделей распределений таков: при существующей точности экспериментальных экологических данных для их описания, как правило, достаточна любая однопараметрическая модель, невысокая точность данных, как правило, не позволяет отдать предпочтение одной из моделей, применение многопараметрических моделей для таких данных нецелесообразно.
Применение распределений
гораздо интереснее, чем подбор аппроксимирующих моделей, другой аспект исследования — применение ранговых распределений для диагностики экологического состояния сообществ на шкале “норма - патология”.
Обнаружено (Левич, 1980), что в нормальном (ненарушенном, фоновом и т.п.) состоянии сообщества параметр рангового распределения заключен во вполне определенном диапазоне значений. Например, для численностей планктонных организмов в случае экспоненциальной модели z » 0,7 -0,9 или в случае гиперболической модели b » 1,5 -2,5. (Напомню, что в лингвистике для “целостных” текстов выполняется b = 1 (Zipf, 1949; Орлов, 1970).) Параметр рангового распределения специфичен для типа сообщества (например, для сообществ фитопланктона, зоопланктона или перифитона), для конкретной экосистемы, для сложившегося комплекса условий среды, к которым адаптировано сообщество. В той степени, в какой справедлив указанный закон, отклонения от него могут служить мерой патологии состояний сообщества. Другими словами, предлагается “градусник” для экосистем, где роль температуры играет параметр рангового распределения (замечу, что приведенная аналогия не является поверхностной метафорой, о чем пойдет речь в разделе о происхождении ранговых распределений).
Укоренившаяся в экологии практика диагностики состояния экосистем по индексам их разнообразия, тем самым, имеет свое оправдание в том, что все индексы разнообразия однозначно связаны с параметрами ранговых распределений (Левич, 1980) и сами параметры чаще всего интерпретируются как индексы экологического разнообразия.
Следует уточнить, что отклонения ранговых распределений от нормы регистрируют стрессовые воздействия на сообщества. При длительном сохранении нарушающего воздействия может произойти существенная перестройка структуры сообщества, замена входящих в него видов, но в результате адаптации параметры рангового распределения новых численностей новых групп организмов окажутся в пределах нормы.
Ранговые распределения применялись для анализа процессов эвтрофирования вод, для оценки влияния на биоту загрязняющих веществ и теплового загрязнения, для изучения сукцессий, сезонных изменений и многого другого (Inagaki and Lenoir, 1974; Bazzaz, 1975; Левич, 1980; Сироткина и Левич, 1981; Lecordier and Lavielle,1982; Сахаров и Ильяш, 1982; Максимов и др., 1996).
Распространен метод диагностики, основанный на сравнении ранговых распределений численностей и биомасс одних и тех же групп организмов (Warwick, 1987). Этот АВС-метод (abundance-biomass comparison) основан на предположении о том, что в стабильных сообществах преобладают крупные виды с медленной динамикой, а в нарушенных — мелкие и более динамичные. Пользователи метода, как правило, предлагают свои способы расчета индексов отличия указанных распределений (Beukema, 1988; McManus and Paulv,1990; Meire and Deren, 1990; Аверинцев, 1991).
На Биологическом факультете МГУ А.Азовским создана компьютерная программа ЭКОС для анализа экологических данных, где, в частности, содержится модуль для исследования видовой структуры сообществ (расчет индексов разнообразия, подбор формы рангового распределения, вычисление параметров этого распределения и их погрешностей, АВС-метод).
Метод оценки экологического состояния природных объектов с помощью параметров ранговых распределений открывает подход к региональному нормированию нарушающих экологическое благополучие воздействий. Суть подхода состоит в дискриминации уровней воздействий по отношению к границе области нормального функционирования экосистем (Левич, 1994; Максимов и др., 1996).
Происхождение распределений
Существующие подходы. Известно несколько подходов к обоснованию формы ранговых распределений. Например, объяснение экспоненциального закона Мотомуры (Motomura, 1932) с помощью априорных представлений а различном “могуществе” видов при взаимодействии в сообществе. Беспараметрическая модель “разломанного стержня” МакАртура (MacArthur, 1960) рассчитывает результат случайного бросания точек на отрезок (“стержень”), олицетворяющий доступный сообществу запас единственного и равномерно распределенного ресурса. В.Д.Федоров (1978) в модели “экспоненциально разломанного стержня” делает то же на отрезке с экспоненциально распределенным ресурсом. Р.Кернер (Песенко, 1978) показал, что логнормальный закон следует из статистической механики системы “хищник-жертва”, описываемой уравнениями Вольтерра. Гиперболическую зависимость числа видов с данным значением существенного параметра от величины этого параметра выводят Б.Я.Рябко, Б.И.Кудрин, Н.Н.Завалишин и А.И.Кудрин (1978). Адекватную форму ранговых распределений для численностей биологических таксонов на основе модели самоподобных множеств выводят Ю.Г.Пузаченко и А.Ю.Пузаченко (1996).
Особое место в обосновании ранговых распределений занимают подходы, основанные на экстремальных принципах. Как правило, это вариации принципа максимума энтропии (Шрейдер, 1967; Мандельброт, 1973; Арапов, Шрейдер, 1977; Яблонский, 1986; Кудрин и др., 1987). Разработана модель (Левич, 1982; Левич и др., 1993; Левич и др., 1994; Levich, 1995), развивающая идеи экстремального подхода для обоснования распределений численности классов сложных открытых систем в условиях конкуренции за ресурсы. От существующих моделей ее отличают, в первую очередь, два обстоятельства. Первое из них — вывод, а не постулирование экстремизируемого функционала, и второе — учет ресурсных ограничений при условной оптимизации в виде неравенств, а не равенств, что в силу большей реалистичности и общности позволяет решать ряд задач, недоступных традиционному подходу.
Категорно-функторная модель сообщества. При формальном описании естественные системы моделируются математическими структурами. (Например, физическое пространство — структурой трехмерного евклидова действительного многообразия; состояния атома — структурой бесконечномерного гильбертова пространства; различные сообщества — структурой множества с разбиениями.) Математические методы позволяют строго сравнивать между собой математические структуры. Предлагается экстремальный принцип: в реальности осуществляются состояния систем, описываемые экстремальными математическими структурами (в пределах доступных системам ресурсов). Для описания математических структур удобен язык теории категорий и функторов (Левич, 1982). Множества без структуры сравниваются при помощи кардинальных чисел (мощностей), эксплицирующих понятие количества элементов. Прямым обобщением кардинальных чисел на структурированные множества являются структурные числа, однако в отличие от кардинальных — структурные числа упорядочены не линейно, а лишь частично. Функторный метод сравнения математических структур позволяет сравнивать их с помощью числовых инвариантов, которые рассчитываются как количество допустимых структурой системы преобразований. Так, например, для структуры множеств с разбиениями, если допустимы “размножение” и “смертность” элементов множества (“особей”) и не допустимы их слияние друг с другом и интродукция извне, а каждый класс элементов переходит в себя же (для экологических сообществ это соответствует сохранению биологических видов), удельный инвариант есть:
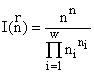 ,
,
где ni — численность класса i; ![]() , w — число классов разбиения (например, видов в сообществе) и
, w — число классов разбиения (например, видов в сообществе) и ![]() обозначает совокупность (n1, n2, ..., nn). Если допустимы как размножение, смертность, так и слияние и интродукция особей, то инвариант структуры множеств с разбиениями в этом случае есть
обозначает совокупность (n1, n2, ..., nn). Если допустимы как размножение, смертность, так и слияние и интродукция особей, то инвариант структуры множеств с разбиениями в этом случае есть
![]()
(другие типы инвариантов приведены в работах Левич, 1982; Levich, 1995).
Рассчитанные инварианты могут теперь играть роль экстремизируемых функций в соответствующих вариационных задачах (технически более удобно работать с их логарифмами). Приведу вариационную задачу для одного из инвариантов:
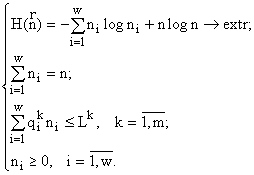
Здесь m — число ресурсов, потребляемых системой; Lk — доступное системе количество ресурса k в среде (Lk >0); ![]() — содержание (потребность) ресурса k в “особи” из класса i
— содержание (потребность) ресурса k в “особи” из класса i ![]() ; искомыми функциями являются численности классов ni и их сумма n.
; искомыми функциями являются численности классов ni и их сумма n.
Показано, что решение этой задачи существует, осуществляет максимум функционала, единственно и выражается “формулой видовой структуры”
![]() ,
,
где величина n и множители Лагранжа lk являются функциями ресурсов ![]() и отыскиваются из системы
и отыскиваются из системы
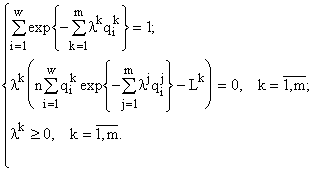
Обращу внимание на ряд особенностей модели. Рассматриваемые функционалы имеют “энтропиеобразный” вид. И это не случайно, так как и при вычислении больцмановской энтропии и при подсчетах инвариантов фактически рассчитывается одна и на же величина — логарифм количества допустимых микросостояний (допустимых преобразований), соответствующих заданному макросостоянию (структуре) системы. Таким образом, инварианты могут рассматриваться как “обобщенные энтропии” математических структур. Существенно, что эти “энтропии” точно рассчитываются вне каких-либо статистических представлений и для любых, сколь угодно малых или больших численностей ni. Многие их полученных инвариантов известны в экологии как императивные (в отличие от параметрических — параметров ранговых распределений) индексы разнообразия. Так, например, приведенные выше инварианты соответствуют индексам МакАртура (MacArthur, 1955) и Симпсона (Simpson, 1949).
Приведу имеющиеся интерпретации предложенного экстремального принципа:
- максимальная по силе структура системы, или состояние, наиболее удаленное от своего бесструктурного аналога;
- наименьшее количество допустимых структурой системы преобразований (т.е. наибольшая устойчивость) при максимальной экспансии системы (росте тотальной численности n);
- наибольшее разнообразие системы (ее сложность, степень самоорганизации или содержание информации в ней (Левич, 1978));
- наименьшее потребление любого из лимитирующих рост системы ресурсов (см. теорему Гиббса в нижеследующих разделах текста).
Формула видовой структуры и ранговые распределения. В формуле видовой структуры номер i маркирует классы сообщества и не обязательно соответствует рангу класса по численности. Предположим, что маркировка выбрана в соответствии с рангами, тогда формула видовой структуры превращается в ранговое распределение численностей.
Пусть m = 1, т.е. потребляется единственный ресурс. Пусть потребности видов в этом ресурсе распределены линейно: -qi º
q1i. Тогда формула видовой структуры задает экспоненциальное ранговое распределение ni/n = zi, где z = e-lq1. Пусть потребности видов распределены по закону qi = q0 ln(B +i), тогда ni/n ~ 1/(B +i)b, где b=lq0. Для такого медленноменяющегося распределения потребностей, как qi ~ lnlni, получаем распределение численностей, близкое к задаваемому моделью “разломанного стержня” распределению ni ~ lni. В случае двух ресурсов и распределений потребностей ![]() и
и ![]() получаем двухпараметрическое дзета-распределение численностей. Становится понятной резко убывающая форма ранговых распределений: согласно формуле видовой структуры, численности экспоненциально зависят от монотонных распределений потребностей. Таким образом, распределение численностей полностью порождается распределением их потребностей. Параметры ранговых распределений пропорциональны множителям Лагранжа соответствующей вариационной задачи. (Замечу, что в задачах статистической физики принцип максимума энтропии использовали неоднократно (Больцман, 1953; Gibbs, 1902, Jaynes, 1957; Haken, 1988). При рассмотрении газа и ограничений по энергии величина 1/l интерпретируется как температура газа T.)
получаем двухпараметрическое дзета-распределение численностей. Становится понятной резко убывающая форма ранговых распределений: согласно формуле видовой структуры, численности экспоненциально зависят от монотонных распределений потребностей. Таким образом, распределение численностей полностью порождается распределением их потребностей. Параметры ранговых распределений пропорциональны множителям Лагранжа соответствующей вариационной задачи. (Замечу, что в задачах статистической физики принцип максимума энтропии использовали неоднократно (Больцман, 1953; Gibbs, 1902, Jaynes, 1957; Haken, 1988). При рассмотрении газа и ограничений по энергии величина 1/l интерпретируется как температура газа T.)
В качестве спекулятивной иллюстрации приведу доводы в пользу ципфовской модели рангового распределения встречаемости слов в текстах. Пусть i обозначает ранг встречаемости некоторого слова в текстах, по которым происходит обучение индивидуума языку. Введем понятие “усилия l(i) по воспроизведению слова i“ при создании текстов. Согласно физиологическому закону Фехнера величины стимула и реакции связаны логарифмической зависимостью. Поэтому “потребность“ qi слова i в усилии по его воспроизведению при создании текста можно считать пропорциональной log i, и, следовательно, встречаемость этого слова в тексте и его ранг окажутся связанными гиперболической зависимостью.
Теоремы вариационного моделирования. Для приведенной выше вариационной задачи доказан ряд теорем, позволяющих решать проблемы управления структурой сообществ.
Теорема стратификации (Левич и др., 1994). Пространство ресурсов среды ![]() разбивается (стратифицируется) на 2m -1 непересекающихся областей SJ
разбивается (стратифицируется) на 2m -1 непересекающихся областей SJ ![]() , в каждом из которых:
, в каждом из которых:
- ресурсы kÎ J и только они потребляются полностью (неравенства вариационной задачи превращаются в строгие равенства, а для ресурсов kÏ J неравенства обращаются в строгие неравенства);
- численности ni (через параметры n и ![]() из формулы видовой структуры) зависят только от величин Lk для ресурсов kÎ
J.
из формулы видовой структуры) зависят только от величин Lk для ресурсов kÎ
J.
Теорема стратификации позволяет по заданным наборам ![]()
![]() определить, в каком страте лежит заданный вектор ресурсов
определить, в каком страте лежит заданный вектор ресурсов ![]() , т.е. какие ресурсы фактически определяют (лимитируют) функционирование сообщества.
, т.е. какие ресурсы фактически определяют (лимитируют) функционирование сообщества.
Теорема оптимизации (Левич и др., 1993). Относительные численности классов ni/n зависят только от отношений Lk/Lj лимитирующих ресурсов; эти численности достигают максимальной величины при отношениях ресурсов, равных отношениям соответствующих потребностей. Теорема оптимизации задает путь управления (стимулирования или подавления) видовыми обилиями ni/n с помощью отношений лимитирующих ресурсов в среде.
Теорема Гиббса (Левич и Алексеев, 1996). Для любого ресурса j рассмотренная вариационная задача на максимум энтропии при ограничениях сверху на потребленные ресурсы эквивалентна следующей вариационной задаче:

где H0 — некоторая константа (однозначно связанная с параметрами исходной задачи). Роль теоремы Гиббса для интерпретации экстремального принципа уже отмечалась.
От биоценозов и экологии к техноценозам и экономике
Решаемые моделью задачи возникают, конечно, не только в экологии, но и в технике (Кудрин, 1993) и в экономике (напомню, что, по Геккелю, экология — это “экономика природы”). Приведу примеры многокомпонентных открытых систем в условиях конкуренции за многочисленные ресурсы:
- Технические системы, состоящие из когорт агрегатов различных классов. Например, система электроснабжения, включающая электродвигатели и другие силовые установки, потребляющие электрическую энергию, эксплуатационные и ремонтные материалы, средства на обновление и ремонт, людские ресурсы и т. д.
- Экономическая система, состоящая из классов предприятий, объединенных по некоторому признаку. Например, предприятия отрасли производства, объединенные в группы идентичных по мощности, размеру, потребностям в ресурсах. Предприятия потребляют сырье, материалы, воду, энергию, инвестиции, кредиты, рабочую силу и т. д.
- Производственная система с многопрофильной продукцией, объединяющая производство нескольких типов товаров и потребляющая общие для этих товаров ресурсы.
Приведу формулировки проблем для описываемых систем. Пусть известны:
- вектор ![]() имеющихся в распоряжении системы ресурсов, подчиняющихся законам сохранения (т.е. не информационных, не поведенческих, не моральных и т.д.);
имеющихся в распоряжении системы ресурсов, подчиняющихся законам сохранения (т.е. не информационных, не поведенческих, не моральных и т.д.);
- вектора ![]() потребностей объектов класса i в ресурсах
потребностей объектов класса i в ресурсах ![]() . Потребности
. Потребности ![]() –системообразующий фактор, т.е. в класс i объединяются все потребители с одинаковыми потребностями
–системообразующий фактор, т.е. в класс i объединяются все потребители с одинаковыми потребностями ![]() . Некоторые из
. Некоторые из ![]() могут быть нулевыми.
могут быть нулевыми.
Пусть необходимо отыскать численности классов ![]() , i =
, i =![]() . (Номер i лишь маркирует класс, т.е. порядок номеров не существенен.)
. (Номер i лишь маркирует класс, т.е. порядок номеров не существенен.)
Вариационное моделирование позволяет ставить и решать следующие задачи для указанного класса систем:
- выявление ресурсов, ограничивающих функционирование системы, т.е. отыскание подмножества ![]() ресурсов такого, что ресурсы
ресурсов такого, что ресурсы ![]() потребляются системой полностью и добавление их в систему приводит к росту численностей
потребляются системой полностью и добавление их в систему приводит к росту численностей ![]() .
.
- расчет функций ![]()
![]() .
.
- расчет парциальных потреблений![]() , лимитирующих ресурсов каждым классом i системы
, лимитирующих ресурсов каждым классом i системы  .
.
- расчет реальных потреблений ![]() и
и ![]() (всей системой и каждым ее классом) ресурсов, не являющихся лимитирующими (эти ресурсы не потребляются из среды полностью), другими словами, расчет сценариев сбалансированного потребления ресурсов.
(всей системой и каждым ее классом) ресурсов, не являющихся лимитирующими (эти ресурсы не потребляются из среды полностью), другими словами, расчет сценариев сбалансированного потребления ресурсов.
- оптимизация численностей классов ![]() , т.е. расчет комбинации параметров задачи
, т.е. расчет комбинации параметров задачи ![]() , при которых численность
, при которых численность ![]() максимальна или минимальна. Другими словами, указание пути управления “видовой” структурной системы.
максимальна или минимальна. Другими словами, указание пути управления “видовой” структурной системы.
- расчет интегральных характеристик системы: ранговых распределений n(j); индексов разнообразия системы и др.
- оценка состояния системы в терминах нормы и патологии с помощью интегральных характеристик. Расчеты допустимых уровней воздействий, нарушающих нормальное функционирование системы.
Список литературы.
Аверинцев В.Г. Оценка сезонной динамики функционального состояния высокоарктических мелководных экосистем Земли Франца-Иосифа методом АВС // Проблемы экологии полярных областей. Вып. 2. М.: Наука. 1991. С. 23-24.
Арапов М.В., Шрейдер Ю.А. Классификация и ранговые распределения // Научно-техническая информация. Сер. 2. 1977. №11-12. С. 15-21.
Больцман Л. Лекции по кинетической теории газов. М.: Гостехиздат. 1953.
Кудрин Б.И. Введение в технетику. Томск: Издательство Томского университета. 1993.
Кудрин Б.И., Кудряшов С.А., Фуфаев В.В., Якимов А.Е. Канонизация и управление видовой структурой ценоза. Принцип максимума энтропии // Доклады МОИП. 1987. Общая биология. Ìîðôîëîãèÿ и генетика процессов роста и развития. М.: Наука. 1989. С.
Левич А.П. Информация как структура систем // Семиотика и информатика. Вып. 10. 1978. С. 116-132.
Левич А.П. структура экологических сообществ. М.: Издательство Московского университета. 1980.
Левич А.П. Теория множеств, язык теории категорий и их применение в теоретической биологии. М.: Издательство Московского университета. 1982.
Левич А.П. Биотическая концепция контроля природной среды // Доклады Академии наук. 1994. Т. 337. №2. С. 257-259.
Левич А.П., Алексеев В.Л., Рыбакова С.Ю. Оптимизация структуры экологических сообществ: модельный анализ // Биофизика. 1993. Т. 38. Вып. 5. С. 877-885.
Левич А.П., Алексеев В.Л., Никулин В.А. Математические аспекты вариационного моделирования в экологии сообществ // Математическое моделирование. 1994. Т. 6. №5. С. 55-71.
Левич А.П., Алексеев В.Л. Энтропийный экстремальный принцип в экологии сообществ: результаты и обсуждение // Биофизика. 1996 (в печати).
Максимов В.Н., Джабруева Л.В., Булгаков Н.Г., Терехин А.Т. Концепция выявления стрессовых состояний водных экосистем методом ранговых распределений и экологически допустимые уровни загрязняющих веществ для водоемов р. Элиста // Водные ресурсы. 1996 (в печати).
Мандельброт Б. Теория информации и психологическая теория частот слов // Математические методы в социальных науках. М.: Наука. 1973. С. 316-447.
Орлов Ю.К. О статистической структуре сообщений, оптимальных для человеческого восприятия // Научно-техническая информация. Сер. 2. 1970. №8.
Песенко Ю.А. Концепция видового разнообразия и индексы его измеряющие // Журнал общей биологии. 1978. Т. 39. №3.С. 380-393.
Пузаченко Ю.Г., Пузаченко А.Ю. Семантические аспекты биоразнообразия // Журнал общей биологии. 1996. Т. 57. №1. С. 1-43.
Рябко Б.Я., Кудрин Б.И., Завалишин Н.Н., Кудрин А.И. Модель формирования статистической структуры биоценозов // Известия СО АН СССР. Сер. биол. 1978. Вып. 1. С. 121-127.
Сахаров В.Б., Ильяш Л.В. Применение метода функции желательности к анализу результатов изучения действия цинка и хрома на фитопланктон Рыбинского водохранилища // Биологические науки. 1982. №8. С. 65-68.
Сироткина Н.В., Левич А.П. Влияние тяжелых металлов на видовую и надвидовую структуры фитопланктонного сообщества Рыбинского водохранилища // Человек и биосфера. Вып. 6. М.: Издательство Московского университета. 1981. С. 142-150.
Федоров В.Д. Относительное обилие симпатрических видов и модель экспоненциально разломанного стержня // Человек и биосфера. Вып. 2. М.: Издательство Московского университета. 1979. С. 17-41.
Шрейдер Ю.А. О возможности теоретического вывода статистических закономерностей текста // Проблемы передачи информации. Вып. 1. М.: Наука. 1967. С. 3-10.
Яблонский А.И. Математические модели в исследовании науки. М.: Наука. 1986.
Bazzaz F.A. Plant species diversity in old-field successional ecosystems in Southern Illinois // Ecology. 1975. V. 56. Pp. 485-488.
Beukema J.J. An evaluation of the ABC-method as applied to macrozoobenthic communities living on tidal flats in the Dutch Wadden Sea // Mar. Biol. 1988. V. 99. Pp. 425-433.
Frontier S. diversity and structure in aquatic ecosystems // Oceanogr. and Mar. Biol. Annual Rev. 1985. V. 23. Pp. 253-278.
Gibbs J.W. Elementary principles in statistical mechanics. N.Y.: Longmans, Green & Co. 1902.
Haken H. Information and Self-Organization. A Macroscopic Approach to Complex Systems. Berlin: Springer-Verlag. 1988.
Inagaki H., Lenoir A. Une etude d’ecologie evolutive: application de la loi de Motomura aux fourmis // Bull. Ecol. 1974. V. 5. ¹3. 207-219.
Jaynes E.T. Information theory and statistical mechanics // Phys. Rev. 1957. V. 106. ¹4. Pp. 620-630; V. 108. ¹2. Pp. 171-190.
Lecordier C., Lavelle P. Application du modele de Motomura aux peuplements de vers de terre: signification et limites // Rev. Ecol. Et Biol. Sol. 1982. V. 19. ¹2. 177-191.
Levich A.P. Time as variability of natural systems: ways of quantitative description of changes and creation of changes by substantial flows // On the Way to Understanding the Time Phenomenon: The Constructions of Time in Natural Science. Singapore: World Scientific. 1995.
MacArthur R.H. Fluctuations of animal populations and measure of community stability // Ecology. 1955. V. 36. ¹7. Pp. 533-536.
MacArthur R.H. On the relative abundance of species // Amer. Nat. 1960. V. 94 Pp. 25-36.
McManus J.W., Paulv D. Measuring ecological stress: variations on theme by R.M. Warwick // Mar. Biol. 1990. V. 108. ¹2. Pp. 305-308.
Meire P.M., Deren J. Use of the abundence-biomass comparison method for detecting environmental stress: some considerations based on intertidal macrozoobenthos and bird communities // J. Appl. Ecol. 1990. V. 27 ¹1. Pp. 210-221.
Motomura I. A statistical treatment of associations // Japan J. Zool. 1932. V. 44. Pp. 379-387 (in Japanese).
Simpson E.H. Measurement of diversity // Nature. 1949. V. 169 P. 688.
Zipf G.K. Human Behaviour and the Principle of Least Effort. An Introduction to Human Ecology. Cambridge: Addison-Wesley Press. 1949.