УДК 577.4:579.841.11.017.6
Потребности диссоциантов Pseudomonas aeruginosa в глюкозе, нитратах и фосфатах. Лимитирующие рост концентрации веществ при культивировании без пополнения запаса ресурсов*
©2001 г. В. Н. Максимов, Е. С. Милько, А. П. Левич
Биологический факультет Московского государственного университета им М.В. Ломоносова
119899, Москва, Воробьевы горы;
Поступила в редакцию 01.2001 г.
По данным 250 опытов при культивировании без пополнения запаса ресурсов R-, S- и М-диссоциантов штамма Pseudomonas aeruginosa К-2 на средах с различным начальным содержанием глюкозы, нитратов и фосфатов рассчитаны потребности диссоциантов в указанных ресурсах и погрешности в определении потребностей. В пределах погрешностей потребности в ресурсах R- и S-диссоциантов оказываются неразличимыми. Потребности М-диссоцианта заметно отличаются от потребностей R- и S-форм, но из-за особенностей роста М-формы доверительные интервалы измеренных величин могут перекрываться с доверительными интервалами форм R и S. Полученные значения потребностей позволяют оценить концентрации исследуемых ресурсов, лимитирующие рост культур.
Ключевые слова: потребности в ресурсах, диссоцианты бактерий, Pseudomonas aeruginosa, принцип лимитирования, лимитирующие концентрации.
ПОНЯТИЕ ПОТРЕБНОСТИ
Цель предлагаемой вниманию экологов и микробиологов работы – получить оценки потребностей в основных компонентах питания для физиологически различающихся популяций микроорганизмов. Количественные данные о потребностях необходимы для предсказания лимитирующих рост концентраций в питательных средах и для анализа конкуренции за ресурсы в лабораторных поликультурах.
Многолетние эксперименты с накопительными культурами одноклеточных организмов показывают, что количество вновь образовавшихся клеток и количество потребленного из среды лимитирующего ресурса оказываются прямо пропорциональными друг другу, если рост культуры ограничен единственным питательным ресурсом. Видоспецифический коэффициент пропорциональности q![]() =
=![]() L/
L/![]() n, имеющий смысл количества ресурса, необходимого для новообразования одной клетки, логично назвать потребностью вида i в ресурсе L. Фактически величина потребности q (в пересчете с численности на биомассу клеток) обратна экономическому коэффициенту, значения которого обычно фигурируют при описании процесса культивирования (Перт, 1978).
n, имеющий смысл количества ресурса, необходимого для новообразования одной клетки, логично назвать потребностью вида i в ресурсе L. Фактически величина потребности q (в пересчете с численности на биомассу клеток) обратна экономическому коэффициенту, значения которого обычно фигурируют при описании процесса культивирования (Перт, 1978).
При измерении потребностей клеток очень важны два обстоятельства. Первое из них – следует оценить, какое количество ресурса из среды идет на увеличение его запаса в клетках и какое - непосредственно на рост. Собственный опыт ведения бактериальных культур (в отличие от культур фитопланктона) показывает, что у бактерий в процессе культивирования не изменяется внутриклеточное содержание таких ресурсов, как углерод, азот, фосфор. Второе обстоятельство – следует быть уверенными, что рост культуры ограничен именно тем единственным ресурсом, потребность в котором измеряется в опыте. В качестве операционального определения лимитирующего ресурса примем правило: компонент питания считается ограничивающим рост культуры, если его добавление в среду после остановки роста приводит к возобновлению деления клеток. Соответственно, если увеличение некоторого ресурса в среде не приводит к росту культуры, можно утверждать, что её рост ограничивают какие-то другие факторы. Если исследователь уверен, что рост культуры ограничен заданным ресурсом, то косвенное измерение потребности клеток в этом ресурсе сводится к прямому измерению двух характеристик культуры – прироста клеток за определенный период времени и убыли исследуемого ресурса из среды за то же время.
МАТЕРИАЛЫ И МЕТОДЫ
Источником данных для исследования послужили более чем 250 опытов по культивированию R-, S- и M-диссоциантов штамма Pseudomonas aeruginosa К-2 на средах с различным начальным содержанием глюкозы, нитратов и фосфатов, проведенные в течение двух лет. (Максимов и соавт., 1999).
Базовая среда для культивирования содержала 3% глюкозы для R- и S-диссоциантов и 1% глюкозы для М-диссоцианта; 1% азотнокислого натрия; 0.15% натрия фосфорнокислого однозамещенного; 0.06% хлористого калия и 0.02% сернокислого магния. Изучение зависимости роста от изменения начальных концентраций ресурсов проводили при варьировании содержания глюкозы в среде от 1 до 7 %, нитратов от 0.01 до 2%, фосфатов – от 0.001 до 0.5%. В каждом диапазоне опыт проводили с 10–12 начальными концентрациями вещества в 2–3-х повторностях. Для контроля потенциальной возможности роста культур на внутриклеточных запасах в каждую серию опытов входили испытания на средах с нулевым содержанием одного из исследуемых питательных веществ.
Бактерии культивировали в пробирках объемом 50 мл с 10мл среды на качалке, обеспечивающей 180 оборотов в минуту, при температуре 30° С. В качестве посевного материала использовали односуточные культуры диссоциантов псевдомонад, выращенных на плотной среде, содержащей в равных отношениях мясо-пептонный бульон и сусло. Бактерии со скошенного агара с помощью микробиологической петли переносили в пробирку с физиологическим раствором. Плотность инокулята выравнивали по нефелометру. Рост бактерий оценивали нефелометрически по плотности культуры. Оптическую плотность переводили в плотность численности культуры с помощью коэффициента: одно деление нефелометра соответствует численности 108 кл/мл. При переводе в единицы численности учтена установленная (Милько, Мартынкина, 1996) неизменность размера клеток Р. aeruginosa в динамике роста культуры.
Данные экспериментов описывали кусочно-линейной моделью:
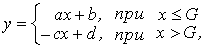
где у - оптическая плотность культур на стационарной стадии роста; х - начальное содержание одного из ресурсов в среде; a, b, c, d и G - параметры модели, которые оценивались методами нелинейного регрессионного анализа с помощью соответствующей программы из пакета STATISTICA 5.O.
Погрешность параметров a, b, c, и d определяли, как ε (z) = ![]() где z – параметр, а Δz – его 95% доверительный интервал: Δz = s{z}tα (f). Стандартное отклонение s{z} рассчитывали отдельно для каждого из линейных участков (т.е. при х<G и при х>G), используя программы линейной регрессии. Табличное значение критерия Стьюдента tα (f) находили для уровня значимости α = 0.05 и числа степеней свободы f = n – 2, где n – число экспериментальных точек на выбранном линейном участке.
где z – параметр, а Δz – его 95% доверительный интервал: Δz = s{z}tα (f). Стандартное отклонение s{z} рассчитывали отдельно для каждого из линейных участков (т.е. при х<G и при х>G), используя программы линейной регрессии. Табличное значение критерия Стьюдента tα (f) находили для уровня значимости α = 0.05 и числа степеней свободы f = n – 2, где n – число экспериментальных точек на выбранном линейном участке.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Не обнаружен рост диссоциантов на внутриклеточных запасах ресурсов в опытах с отсутствием ресурса в среде. (Заметим, что при культивировании фитопланктона (Левич и соавт., 1997) такие запасы обнаружены.)
На рис. 1, где приведены опытные и модельные зависимости плотности культуры от начальной концентрации ресурсов в среде, отчетливо видны границы, в пределах которых каждый из ресурсов является лимитирующим: от нулевой концентрации до точки “перелома” G, после которой увеличение начального содержания питательного вещества в среде не приводит к возрастанию плотности культуры. В пределах лимитирования доступный культуре из среды ресурс и плотность культуры прямо пропорциональны друг другу (коэффициент множественной корреляции между опытным данными и линейной моделью составляет 0.88–0.99).
В табл. 1 приведены значения параметров a, b, c, d и G модели для всех диссоциантов i и ресурсов L. Коэффициент пропорциональности между доступным ресурсом и приростом плотности культуры, обратный параметру а![]() модели, как раз и представляет собой потребность q
модели, как раз и представляет собой потребность q![]() диссоцианта i в ресурсе L. В таблице 2 сведены значения потребностей в единицах потребленного вещества в расчете на одну клетку прироста культуры. Свободные члены в “восходящих” прямых модели малы и в некоторых случаях значимо не отличаются от нуля (их погрешность близка к 100%). Исключение составляют опыты с потреблением фосфатов. Это может означать, что при низких (порядка 10-3 %) концентрациях фосфатов в питательной среде существенное влияние оказывают добавки фосфатов, попадающие в питательную среду вместе с инокулятом.
диссоцианта i в ресурсе L. В таблице 2 сведены значения потребностей в единицах потребленного вещества в расчете на одну клетку прироста культуры. Свободные члены в “восходящих” прямых модели малы и в некоторых случаях значимо не отличаются от нуля (их погрешность близка к 100%). Исключение составляют опыты с потреблением фосфатов. Это может означать, что при низких (порядка 10-3 %) концентрациях фосфатов в питательной среде существенное влияние оказывают добавки фосфатов, попадающие в питательную среду вместе с инокулятом.
“Горизонтальный” участок зависимостей плотности культур от начальных концентраций ресурсов в среде соответствует области концентраций, которые не являются лимитирующими для исследованных монокультур. Степень “горизонтальности” может быть оценена близостью к нулю параметра c модели (табл.1). Положительные значения параметра c соответствуют возможному ингибирующему действию повышенных концентраций ресурсов. Значения параметра c, как правило, или близки к нулю или незначимы при принятом 95% уровне доверительной вероятности. О возможном ингибирующем действии повышенных концентраций ресурсов можно говорить, пожалуй, лишь для действия фосфатов на R-диссоциант и нитратов – на S-диссоциант.
Параметры G![]() (табл.1) использованной в работе регрессионной модели представляют собой границу значения начальных концентраций ресурса L в среде, при превышении которой этот ресурс в условиях проведенных экспериментов уже не ограничивает рост диссоцианта i.
(табл.1) использованной в работе регрессионной модели представляют собой границу значения начальных концентраций ресурса L в среде, при превышении которой этот ресурс в условиях проведенных экспериментов уже не ограничивает рост диссоцианта i.
Отметим некоторые особенности проведенных экспериментов и их интерпретации. Неявно подразумевалось, что остановка роста культур происходила из-за исчерпания в среде лимитирующего ресурса. Это предположение – лишь гипотеза, поскольку химический контроль состава среды не производили. Рост культур M-диссоцианта сопровождался сильным подкислением среды, поэтому, для M-диссоцианта остановка роста с большей вероятностью могла происходить по причинам, не связанным с истощением ресурсов.
ПОТРЕБНОСТИ В РЕСУРСАХ И ЛИМИТИРУЮЩИЕ КОНЦЕНТРАЦИИ РЕСУРСОВ
Области излома графиков на рис.1, или параметры G модели, как указывалось, соответствуют концентрациям исследуемых ресурсов, при которых происходит переключение факторов, лимитирующих рост культуры.
Имеющиеся данные не дают прямых указаний на то, какой именно фактор среды начинает ограничивать рост культуры. Можно предположить, что лимитирование переключается на один из ресурсов, который не варьировали при постановке экспериментов. По полученным значениям потребностей (табл. 2) можно оценить уровень плотности культуры, равный ![]() (где L содержание ресурса в среде, а
(где L содержание ресурса в среде, а ![]() потребность в этом ресурсе диссоцианта i), которого она может достичь по мере исчерпания всего ресурса среды при условии, что все другие факторы среды рост не ограничивают. Проведенные расчеты (табл. 3) показывают, что эти уровни гораздо выше тех, которые реально достигались культурами диссоциантов в проведенных нами опытах,– реально достигнутые уровни примерно соответствуют значениям параметра d модели (табл. 1). Заметим, что уровни d для каждого диссоцианта значимо не отличаются при переключениях лимитирования с различных ресурсов. Этот факт заставляет предполагать, что во всех трех случаях (опыты с вариацией глюкозы, нитратов и фосфатов) причиной переключения становится ограничение роста каким-либо одним и тем же фактором. По нашему мнению, этим фактором является ограничение в потреблении кислорода, обусловленное имевшимся в опыте объемом пробирок и условиями барботажа. Таким образом, оцененные в табл. 1 значения
потребность в этом ресурсе диссоцианта i), которого она может достичь по мере исчерпания всего ресурса среды при условии, что все другие факторы среды рост не ограничивают. Проведенные расчеты (табл. 3) показывают, что эти уровни гораздо выше тех, которые реально достигались культурами диссоциантов в проведенных нами опытах,– реально достигнутые уровни примерно соответствуют значениям параметра d модели (табл. 1). Заметим, что уровни d для каждого диссоцианта значимо не отличаются при переключениях лимитирования с различных ресурсов. Этот факт заставляет предполагать, что во всех трех случаях (опыты с вариацией глюкозы, нитратов и фосфатов) причиной переключения становится ограничение роста каким-либо одним и тем же фактором. По нашему мнению, этим фактором является ограничение в потреблении кислорода, обусловленное имевшимся в опыте объемом пробирок и условиями барботажа. Таким образом, оцененные в табл. 1 значения ![]() обязаны переключению лимитирования глюкозой, или нитратом, или фосфатом на лимитирование кислородом (или еще каким-либо фактором, не связанным с углеродным, азотным или фосфорным питанием).
обязаны переключению лимитирования глюкозой, или нитратом, или фосфатом на лимитирование кислородом (или еще каким-либо фактором, не связанным с углеродным, азотным или фосфорным питанием).
Конкретные значения параметра G определяются как физиологическими характеристиками диссоцианта, так и конкретными начальными значениями не менявшихся в опыте факторов. Чтобы экспериментально найти лимитирующую концентрацию для каких-либо иных значений постоянных факторов (питательных ресурсов, обеспеченности кислородом, кислотности среды, температуры и др.), необходимо провести новые серии экспериментов или обратиться к модельным расчетам. Существуют модели процессов потребления ресурсов и роста в культурах, позволяющие рассчитывать области лимитирования в пространстве ресурсов, если известны потребности в исследуемых ресурсах для клеток в культуре (Левич и соавт., 1993; Левич и соавт., 1997).
Рассмотрим рост монокультуры в случае ограничения роста двумя возможными факторами L1 и L2 (верхние индексы здесь, как и прежде, обозначают не показатели степени, а номер потребляемого ресурса, в то время как нижние индексы ответственны за обозначение типа диссоцианта). Этот случай максимально нагляден. В более сложных (и менее наглядных) случаях необходимые расчеты могут быть проведены и для произвольного числа ресурсов, и для сообщества, включающего произвольное число “видов”. Рис. 2 иллюстрирует правило отыскания лимитирующего ресурса. Прямая q, определяемая вектором ![]() делит пространство ресурсов на две части. При
делит пространство ресурсов на две части. При ![]() рост культуры ограничен ресурсом 1, при
рост культуры ограничен ресурсом 1, при ![]() рост лимитируется ресурсом 2. При
рост лимитируется ресурсом 2. При ![]() оба ресурса ограничивают рост и при пересечении прямой q происходит переключение лимитирования. Абсцисса точки пересечения, например, прямой L1=const с прямой q дает концентрацию G2, при которой происходит переключения лимитирования роста между ресурсами 1 и 2.
оба ресурса ограничивают рост и при пересечении прямой q происходит переключение лимитирования. Абсцисса точки пересечения, например, прямой L1=const с прямой q дает концентрацию G2, при которой происходит переключения лимитирования роста между ресурсами 1 и 2.
В случае трех потенциально ограничивающих рост ресурсов (при этом предполагается, что все другие существенные для роста факторы находятся в избытке) прямая q порождается вектором ![]() в трехмерном пространстве ресурсов. Это пространство делится на 3 области лимитирования участками трех плоскостей, проходящими через прямую q и координатные оси ресурсов. В каждой из этих областей рост культуры ограничен одним из трех ресурсов, а именно тем, координатная ось которого не лежит в данной области. Чтобы определить концентрацию ресурса, при которой происходит переключение лимитирования в опытах, где два другие ресурса постоянны, следует найти соответствующую координату точки пересечения прямой, параллельной изменяемому ресурсу, с одной из трех полуплоскостей, определяющих область лимитирования. Соответствующие расчеты были проведены для векторов потребностей из табл. 2 и данных о не менявшихся в проведенных опытах начальных концентрациях ресурсов. Рассчитанные величины
в трехмерном пространстве ресурсов. Это пространство делится на 3 области лимитирования участками трех плоскостей, проходящими через прямую q и координатные оси ресурсов. В каждой из этих областей рост культуры ограничен одним из трех ресурсов, а именно тем, координатная ось которого не лежит в данной области. Чтобы определить концентрацию ресурса, при которой происходит переключение лимитирования в опытах, где два другие ресурса постоянны, следует найти соответствующую координату точки пересечения прямой, параллельной изменяемому ресурсу, с одной из трех полуплоскостей, определяющих область лимитирования. Соответствующие расчеты были проведены для векторов потребностей из табл. 2 и данных о не менявшихся в проведенных опытах начальных концентрациях ресурсов. Рассчитанные величины ![]() приведены в табл. 4.
приведены в табл. 4.
Чтобы в рамках описанной модели выявить компонент питания, лимитирующий рост монокультуры, для заданных начальных концентраций m ресурсов ![]() , следует установить, в какую область пространства ресурсов попадает точка
, следует установить, в какую область пространства ресурсов попадает точка ![]() . Например, при m = 2 точка А на рис. 2 попадает в область ниже прямой, проходящей через вектор потребностей
. Например, при m = 2 точка А на рис. 2 попадает в область ниже прямой, проходящей через вектор потребностей ![]() . В этой области рост культуры ограничен ресурсом 1. Аналогичные расчеты (Левич и соавт., 1993) для R-диссоцианта Р. aeruginosa, вектор потребностей которого
. В этой области рост культуры ограничен ресурсом 1. Аналогичные расчеты (Левич и соавт., 1993) для R-диссоцианта Р. aeruginosa, вектор потребностей которого ![]() взят из табл. 2, демонстрируют что базовая среда, содержащая 30 мг/мл глюкозы, 10 мг/мл нитрата и 1.5 мг/мл фосфата, принадлежит многогранному углу в пространстве ресурсов, где рост культуры ограничен глюкозой. Для S-диссоцианта (
взят из табл. 2, демонстрируют что базовая среда, содержащая 30 мг/мл глюкозы, 10 мг/мл нитрата и 1.5 мг/мл фосфата, принадлежит многогранному углу в пространстве ресурсов, где рост культуры ограничен глюкозой. Для S-диссоцианта (![]() ) рост ограничен также глюкозой. И для М-диссоцианта (
) рост ограничен также глюкозой. И для М-диссоцианта (![]() ,
, ![]() = {10мг глюкозы/мл; 10 мг нитрата/мл; 1.5 мг фосфата/мл}) рост ограничен глюкозой. Аналогичные результаты о лимитировании компонентов базовой среды дает табл. 3 – именно за счет глюкозы культура достигает наименьшей плотности (правило лимитирования Либиха).
= {10мг глюкозы/мл; 10 мг нитрата/мл; 1.5 мг фосфата/мл}) рост ограничен глюкозой. Аналогичные результаты о лимитировании компонентов базовой среды дает табл. 3 – именно за счет глюкозы культура достигает наименьшей плотности (правило лимитирования Либиха).
ВЫВОДЫ И ОБСУЖДЕНИЕ
Потребности в ресурсах R- и S-диссоциантов в пределах обнаруженных погрешностей (табл. 2) оказываются неразличимыми. Потребности M-диссоцианта заметно отличаются от потребностей R- и S-форм, но из-за особенностей роста M-формы доверительные интервалы измеренных величин велики и могут перекрываться с доверительными интервалами форм R и S. Однако полученные оценки потребностей и лимитирующих концентраций могут служить ориентирами для последующих более строгих измерений. В этих измерениях следует проводить химический анализ среды, прямую проверку причин остановок роста культур, дифференцированную калибровку показаний нефелометра для каждого из диссоциантов, постановку достаточного числа повторностей для снижения величин погрешностей. Постановка опытов должна обеспечить переключение лимитирования с исследуемых ресурсов среды на другие исследуемые ресурсы той же среды, а не на иные факторы (в данной работе таким фактором оказался предположительно кислород). Полученные в работе значения потребностей q и уровни плотности, при которых происходит переключение на лимитирование новым фактором (параметр модели d), позволяют оценить необходимые концентрации исследуемых ресурсов в базовой среде по формуле ![]() (табл. 5).
(табл. 5).
Заметим, что для расчетов, в которых фигурируют потребности клеток, важны не абсолютные величины потребностей, а их отношение между собой, что позволяет не заботиться об единицах измерения потребностей, если необходимые отношения сохраняются. Точно так же для расчета лимитирующих концентраций и управления структурой сообществ микроорганизмов важны не абсолютные концентрации ресурсов в средах, а их отношения между собой.
Если для набора из w монокультур известна матрица ![]() их потребностей в m ресурсах, то такая информация в рамках ряда моделей экологических сообществ (Левич и соавт., 1994; Левич и соавт., 1997) позволит:
их потребностей в m ресурсах, то такая информация в рамках ряда моделей экологических сообществ (Левич и соавт., 1994; Левич и соавт., 1997) позволит:
СПИСОК ЛИТЕРАТУРЫ
Левич А.П., Алексеев В.Л., Никулин В.А. Математические аспекты вариационного моделирования в экологии сообществ // Математическое моделирование. 1994. Т. 6. № 5. С. 55-71.
Левич А.П., Замолодчиков Д.Г., Алексеев В.Л. Правило лимитирующего звена для многовидовых экологических сообществ // Журнал общей биологии. 1993. Т. 54. № 3. С. 282-297.
Левич А.П., Максимов В.Н., Булгаков Н.Г. Теоретическая и экспериментальная экология фитопланктона. М.: Издательство НИЛ, 1997. 192с.
Максимов В.Н., Милько Е.С., Ильиных И.А. Влияние углеродного, азотного и фосфорного питания на рост R-, S- и M- диссоциантов Pseudomonas aeruginosa // Микробиология. 1999. Т. 68. № 2. С. 206-210.
Милько Е.С., Мартынкина Л.П. Морфологические и физиолого-биохимические особенности диссоциантов Pseudomonas aeruginosa // Микробиология. 1996. Т. 65. № 5. С. 352-356.
Перт С.Дж. Основы культивирования микроорганизмов и клеток. М.: Мир, 1978. 331с.
ТАБЛИЦЫ
Таблица 1. Параметры кусочно-линейной регрессионной модели (обозначения параметров в тексте, единицы измерения – на рис. 1) и их относительные погрешности. C- глюкоза, N- нитрат, P- фосфат
|
Диссо- |
Ре- |
Параметры и их относительные погрешности |
|||||||||
|
Циант |
сурс |
a |
e (a) |
b |
e (b) |
c |
e (c) |
d |
e (d) |
G |
e (G) |
|
C |
93.5 |
7 |
-2.8 |
123 |
2.8 |
81 |
129 |
8 |
1.37 |
8 |
|
|
R |
N |
471 |
36 |
5.4 |
177 |
7.8 |
61 |
149 |
5 |
0.28 |
36 |
|
P |
5014 |
34 |
37 |
44 |
40.2 |
45 |
143 |
3 |
0.02 |
34 |
|
|
C |
89.8 |
6 |
-3.4 |
76 |
3.6 |
56 |
131 |
8 |
1.47 |
8 |
|
|
S |
N |
353 |
27 |
9.1 |
58 |
14.9 |
26 |
134 |
4 |
0.34 |
27 |
|
P |
4306 |
27 |
24.9 |
52 |
0.9 |
1344 |
117 |
2 |
0.02 |
27 |
|
|
C |
34.5 |
55 |
5.9 |
112 |
2.2 |
59 |
29.7 |
23 |
0.69 |
55 |
|
|
M |
N |
166 |
44 |
13.2 |
31 |
9 |
28 |
38.3 |
9 |
0.13 |
44 |
|
P |
2500 |
49 |
13 |
37 |
8.6 |
265 |
26.1 |
9 |
0.004 |
49 |
|
Таблица 2. Потребности q диссоциантов P. aeruginosa в ресурсах среды (10-10 мг ресурса на клетку) и их относительные погрешности e (%)
|
Диссоци- |
Глюкоза |
Нитрат |
Фосфат |
|||||
|
ант |
q |
e (q) |
q |
e (q) |
q |
e (q) |
||
|
R |
11 |
7 |
2.1 |
36 |
0.20 |
34 |
||
|
S |
11 |
6 |
2.8 |
27 |
0.23 |
27 |
||
|
M |
29 |
55 |
6.0 |
44 |
0.40 |
49 |
||
Таблица 3. Уровни оптической плотности (показания нефелометра x100, см. рис. 1), достигаемые культурами диссоциантов на базовой среде (3% или 1% глюкозы, 1% нитратов, 0.15% фосфатов) при условии ограничения роста культуры одним из ресурсов питательной среды
|
Лимитирующий ресурс |
|||
|
Диссоциант |
Глюкоза |
Нитрат |
Фосфат |
|
R |
280.5 |
471 |
752 |
|
S |
269.4 |
353 |
646 |
|
M |
34.5 |
166 |
375 |
Таблица 4. Рассчитанные значения ![]() концентраций ресурса L, при которых происходит переключение лимитирования в случае ограничения роста культур i или глюкозой, или нитратом, или фосфатом базовой среды (3% или 1% глюкозы, 1% нитрата, 0.15% фосфата)
концентраций ресурса L, при которых происходит переключение лимитирования в случае ограничения роста культур i или глюкозой, или нитратом, или фосфатом базовой среды (3% или 1% глюкозы, 1% нитрата, 0.15% фосфата)
|
Концентрация ресурса, % |
|||
|
Диссоциант |
Глюкоза |
Нитрат |
Фосфат |
|
R |
5.2 |
0.57 |
0.055 |
|
S |
3.9 |
0.76 |
0.063 |
|
M |
4.8 |
0.21 |
0.014 |
Таблица 5. Наибольшие значения концентраций глюкозы, нитрата и фосфата в питательной среде, при которых ограничение роста диссоциантов возможно только каким-либо из указанных ресурсов
|
Содержание ресурсов в среде (%) |
|||
|
Диссоциант |
Глюкоза |
Нитрат |
Фосфат |
|
R |
1.4 |
0.32 |
0.029 |
|
S |
1.5 |
0.40 |
0.027 |
|
M |
0.86 |
0.23 |
0.010 |