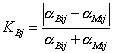Феномен и ноумен времени. Том 1 (1), 2004. С. 50-54.
© Н. Н. Мушкамбаров, Ю. А. Ершов, 1992
Гипотеза об обратимости старения в линии половых клетокН. Н. Мушкамбаров, Ю. А. Ершов |
Московская медицинская академия им. И. М. Сеченова Наличие возрастных изменений с достоверностью установлено практически для всех видов соматических тканей. Но до сих пор не только открытым, но и фактически не поднимавшимся, является вопрос об отношении к старению линии половых клеток. Хотя именно здесь мы можем столкнуться с феноменом, который мог бы иметь принципиальнейшее значение. Речь идёт о том, что старение в линии этих клеток, возможно, является обратимым, и это обращение накопленных прежде возрастных изменений совершается на строго определённом этапе развития данной линий клеток. Для определённости далее мы будем говорить, в основном, о мужских половых клетках, хотя ключевые моменты и выводы справедливы также для женских клеток. Жизненный цикл сперматогенных клетокЧтобы сделать более понятным предмет разговора, предварительно кратко очертим жизненный цикл линии мужских половых клеток. Примечательно, что он характеризуется строгой временной и пространственной упорядоченностью. Первичные половые клетки обособляются на очень ранней (ещё досомитной) стадии развития зародыша в ткани желточного мешка, а во время гаструляции (у человека - к концу первого месяца развития) перемещаются в зачатки гонад. Здесь под влиянием соматических клеток происходит их половая дифференциация - образуются т. н. просперматогонии. На третьем месяце внутриутробной жизни последние многократно интенсивно делятся, а с 4-го месяца наступает длительный период покоя. Вскоре после рождения деление половых клеток на некоторый срок возобновляется, дочерние же клетки (т. н. изолированные сперматогонии, иСг) вновь впадают в "спячку". Этот (уже второй) период покоя продолжается до наступления половой зрелости организма и дальше. Может пройти несколько десятков лет, прежде чем та или иная группа иСг вступит на путь необратимой дифференцировки. Вероятно, запас иСг пополняется за счёт редких спорадических делений. Началом сперматогенеза считают такое деление иСг, после которого между дочерними клетками остаётся цитоплазматический мостик. И все последующие деления - тоже не совсем полные. В итоге, всё потомство одной стволовой клетки, вступившей в дифференцировку (а это, в конечном счёте, несколько тысяч клеток), развивается в виде цитоплазматического синцития. В сперматогенезе - 4 этапа: сперматогоний, сперматоцитов, сперматид и сперматозоидов. На стадии сперматогоний главное - серия митотических делений, с которыми последовательно меняется и морфология клеток. Сперматоциты - это клетки, проходящие ключевой, наиважнейший процесс сперматогенеза - мейоз, т. е. такое деление, которое никаким более клеткам организма не свойственно. В ходе длительной профазы мейоза (которая длится у человека около месяца) гомологичные хромосомы конъюгируют друг с другом. При этом резко активизируются ферменты, разрывающие и сшивающие ДНК. С их помощью хромосомы обмениваются фрагментами ДНК (кроссинговер). И лишь затем происходят два быстрых деления, в результате которых образуются сперматиды, содержащие в ядрах гаплоидный (половинный) набор хромосом. Сперматиды уже более не делятся, и в течение последующего месяца в них происходят сложные морфологические изменения, превращающие их в сперматозоиды. Весь процесс сперматогенеза занимает у человека 75 дней. Но сперматозоиды ещё две-три недели дозревают в придатке семенника. Варианты изменения биологического возраста в линии половых клетокТеперь попытаемся представить, как меняется биологический возраст в обсуждаемой линии клеток. Конечно, "биологический возраст" – понятие не очень определённое. Будем считать, что биологический возраст G - это интегральный показатель, относящийся к каждой отдельной клетке и объединяющий неким образом все характерные признаки её старения. В частности, на уровне ДНК это такие признаки, как разрывы цепей, сшивки цепей друг с другом и с соседними белками, снижение числа метильных групп и т. д. Известны также другие признаки старения на уровне клеток - повреждение мембран, снижение активности тех или иных Ферментов и пр. Не детализируя, насколько конкретно эти признаки увеличивают биологический возраст клетки, естественно считать, что они всё-таки его увеличивают. Тогда G - функция двух параметров:
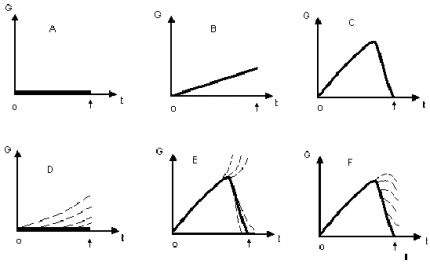
где t - физическое время, а i - номер половой клетки в популяции однородных клеток (нумерация, конечно, произвольная). И вся проблема заключается в том, каков вид функции Мы можем предложить всего шесть вариантов поведения этой функции (см. рис.1). При этом мы рассматриваем интервал времени от начала развития зиготы одного поколения до начала развития зиготы следующего поколения, образовавшейся с участием половой клетки, развившейся из первой зиготы. Следовательно, этот временной интервал включает эмбриональный период развития из зиготы половых клеток, период покоя изолированных сперматогоний (который может длиться от 15 до 50-70 лет), а также период созревания некоторых иСг в сперматозоиды. Первые три варианта предполагают, что G не зависит от i, т.е. все клетки одной стадии развития одинаковы. Вариант А: биологический возраст линии половых клеток на всех стадиях равен нулю. Примерно так считал немецкий ученый Вирхов, когда в начале века выдвинул идею о нестареющей зародышевой плазме. Но это мало согласуется с известными данными о гибели на разных стадиях сперматогенеза и (оогенеза) немалого числа половых клеток. Вариант В: биологический возраст линии половых клеток монотонно увеличивается. Это графическое изображение версии о том, что половые клетки тоже стареют. Но если продолжить график на последующие поколения, получится неуклонный рост. Тогда биологический возраст зиготы каждого очередного поколения будет больше, чем у зиготы предыдущего поколения. С какой бы малой скоростью не происходил этот подъем, в конце концов данный биологический вид должен деградировать и вымирать. Так что это мало правдоподобно. Вариант С – это компромисс между первыми двумя вариантами: клетки стареют, но на какой-то стадии их развития возрастные изменения снимаются – функция Однако еще более вероятны статистические варианты, учитывающие, что старение – процесс вероятностный и однородные клетки стареют по-разному. Тогда вариант А трансформируется в вариант D: клетки стареют, но в течение репродуктивного периода жизни некоторая, все уменьшающаяся доля клеток остается свободной от возрастных изменений. Она-то, с помощью некоторого механизма, и отбирается для оплодотворения. Варианты Е и F – модифицированные варианты С, т.е. варианты, предполагающие обратимость старения. В варианте Е одни клетки, грубо говоря, омолаживаются засчет других (например, передавая им некачественный генетический материал в обмен на нормальные “куски” ДНК). В варианте же F во всех клетках включается механизм “омоложения”, но не везде он снижает G до нуля. Аргументы в пользу “обратимых” вариантов.Таким образом, реальной альтернативой вариантам, предполагающим обратимость старения в линии половых клеток, является лишь вариант D. Но есть группа фактов, свидетельствующих в пользу обратимости. а)Во-первых, почти те же самые ферменты, которые участвуют в кроссинговере, входят и в состав системы репарации ДНК, функционирующей во многих клетках. Эта система выискивает в ДНК места повреждений, вырезает их и синтезирует взамен нормальные фрагменты. Но, очевидно, эта система функционирует не со 100%-ной эффективностью, поскольку повреждения в ДНК с возрастом всё же накапливаются. Можно думать, что это и есть программа старения - постепенное ослабление деятельности системы репарации. В половых же клетках, в процессе профазы мейоза, сходная совокупность ферментов резко активизируется, что в дальнейшем приводит и к снижению количества повреждённых участков. А последующий обмен хромосом участками ДНК может завершить процесс "очистки" определённой части клеток от геномных повреждений. б) Во-вторых, инфузории могут размножаться и половым, и бесполым путём. И, оказывается, половое деление ядер (мейоз) и последующий половой процесс (слияние ядер) резко снижает возраст клеток - культура "молодеет": в ней активизируется обмен веществ, клетки начинают чаще делиться. в) В 1978-1981 гг. Р.Р.Мушкамбаров (с помощью Н.П.Волковой) сравнивал у животных двух возрастов половые клетки одинаковых стадий развития. Для этого из сперматогенных канальцев получали суспензию клеток; последние разделяли специальным методом на 5 фракций, различающихся по степени зрелости. В каждой фракции определяли активность ряда ферментов и содержание белковых фракций (всего - около 25 параметров). После этого рассчитывали коэффициент возрастных различий:
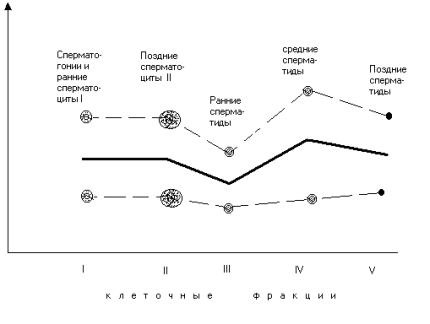
где Оказалось, что возрастные различия (даже по исследованным, достаточно грубым, параметрам) между однородными клетками достоверно существуют. Но в середине сперматогенного ряда, в клетках, только что миновавших мейоз, различия становятся меньше.(рис. 2). (Последующее возрастание степени различий можно объяснить тем, что первые генерации половых клеток от молодых животных, достигшие к моменту эксперимента стадии поздних сперматид и сперматозоидов, являются в биохимическом отношении недостаточно зрелыми. Это было показано на примере одного Фермента - сорбитолдегидрогеназы). РезюмеНа основании сказанного, мы считаем достаточно вероятным предположение, сформулированное в начале доклада, а именно: старение в линии половых клеток обратимо, и это обращение совершается в ходе мейоза.
Рис.№1. Биологический возраст половых клеток: возможные варианты. Ноль и стрелка отмечают моменты образования зигот первого второго поколений.
Рис.№2. Возрастные различия между клетками старых и молодых животных
|